Test: Światło, temperatura i wilgotność jako czynniki środowiskowe. Czynniki fizyczne
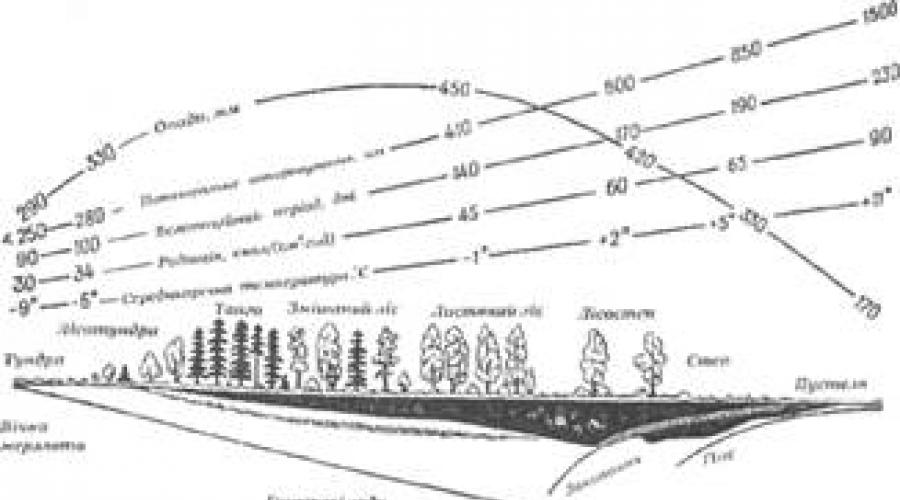
Abiotyczne czynniki środowiskowe obejmują podłoże i jego skład, wilgotność, światło i inne rodzaje promieniowania w przyrodzie oraz jego skład i mikroklimat. Należy zauważyć, że temperaturę, skład powietrza, wilgotność i światło można warunkowo określić jako „indywidualne”, a podłoże, klimat, mikroklimat itp. – do czynników „złożonych”.
Podłoże (dosłownie) to miejsce przywiązania. Na przykład w przypadku roślin drzewiastych i zielnych, w przypadku mikroorganizmów glebowych jest to gleba. W niektórych przypadkach podłoże można uznać za synonim siedliska (np. gleba jest siedliskiem edaficznym). Podłoże charakteryzuje się pewnym składem chemicznym wpływającym na organizmy. Jeśli podłoże rozumieć jako siedlisko, to w tym przypadku jest to zespół charakterystycznych dla niego czynników biotycznych i abiotycznych, do których adaptuje się jeden lub drugi organizm.
Charakterystyka temperatury jako abiotycznego czynnika środowiskowego
Temperatura to czynnik środowiskowy związany ze średnią energią kinetyczną cząstek i wyrażony w stopniach o różnej skali. Najbardziej powszechna jest skala w stopniach Celsjusza (°C), która opiera się na stopniu ekspansji wody (temperatura wrzenia wody wynosi 100°C). W SI przyjmuje się bezwzględną skalę temperatury, dla której temperatura wrzenia wody wynosi T kip. woda = 373 K.
Bardzo często temperatura jest czynnikiem ograniczającym, który decyduje o możliwości (niemożliwości) istnienia organizmów żywych w danym siedlisku.
Ze względu na charakter temperatury ciała a wszystkie organizmy dzielą się na dwie grupy: poikilotermiczne (temperatura ich ciała zależy od temperatury otoczenia i jest praktycznie taka sama jak temperatura otoczenia) i homoitermiczna (temperatura ich ciała nie zależy od temperatury otoczenia i jest mniej więcej stały: jeśli się waha, to w małych granicach - ułamki stopnia).
Poikilothermy obejmują organizmy roślinne, bakterie, wirusy, grzyby, zwierzęta jednokomórkowe, a także zwierzęta o stosunkowo niskim poziomie organizacji (ryby, stawonogi itp.).
Domotermy obejmują ptaki i ssaki, w tym ludzi. Stała temperatura ciała zmniejsza zależność organizmów od temperatury środowiska zewnętrznego, umożliwia zasiedlanie większej liczby nisz ekologicznych, zarówno w rozmieszczeniu równoleżnikowym, jak i pionowym wokół planety. Jednak oprócz homoiotermii organizmy rozwijają adaptacje, aby przezwyciężyć skutki niskich temperatur.
Zgodnie z naturą przenoszenia niskich temperatur rośliny dzielą się na ciepłolubne i odporne na zimno. Do roślin ciepłolubnych należą rośliny południa (banany, palmy, południowe odmiany jabłoni, gruszki, brzoskwinie, winogrona itp.). Rośliny odporne na zimno obejmują rośliny ze średnich i północnych szerokości geograficznych, a także rośliny rosnące wysoko w górach (na przykład mchy, porosty, sosna, świerk, jodła, żyto itp.). W centralnej Rosji uprawia się odmiany odpornych na mróz drzew owocowych, które są specjalnie hodowane przez hodowców. Pierwsze wielkie sukcesy w tej dziedzinie odnieśli I.V. Michurin i inni hodowcy ludowi.
Norma reakcji organizmu na czynnik temperaturowy (dla poszczególnych organizmów) jest często wąska, tj. określony organizm może normalnie funkcjonować w dość wąskim zakresie temperatur. Tak więc kręgowce morskie giną, gdy temperatura wzrasta do 30-32°C. Ale dla żywej materii jako całości granice efektu temperaturowego, w którym życie jest zachowane, są bardzo szerokie. Tak więc w Kalifornii gatunek ryby żyje w gorących źródłach, normalnie funkcjonując w temperaturze 52 ° C, a żaroodporne bakterie żyjące w gejzerach mogą wytrzymać temperatury do 80 ° C (jest to „normalna” temperatura dla ich). W lodowcach w temperaturze -44 ° C niektórzy żyją itp.
Rola temperatury jako czynnika środowiskowego polega na tym, że wpływa ona na metabolizm: kiedy niskie temperatury tempo reakcji bioorganicznych znacznie spowalnia, a przy dużych szybkościach znacznie wzrasta, co prowadzi do zachwiania równowagi w przebiegu procesów biochemicznych, a to powoduje różne choroby a czasem śmierć.
Wpływ temperatury na organizmy roślinne
Temperatura jest nie tylko czynnikiem decydującym o możliwości bytowania roślin na określonym terenie, ale dla niektórych roślin wpływa na proces ich rozwoju. Tak więc ozime odmiany pszenicy i żyta, które podczas kiełkowania nie przeszły procesu „jernalizacji” (niskich temperatur), nie wytwarzają nasion, gdy rosną w najkorzystniejszych warunkach.
Aby wytrzymać działanie niskich temperatur, rośliny mają różne oprawy.
1 w okres zimowy cytoplazma traci wodę i gromadzi substancje działające przeciw zamarzaniu (są to monosacharydy, gliceryna i inne substancje) - stężone roztwory takich substancji zamarzają tylko w niskich temperaturach.
2. Przejście roślin do stadium (fazy) odpornej na niskie temperatury - stadium zarodników, nasion, bulw, cebul, kłączy, roślin okopowych itp. Drzewiaste i krzewiaste formy roślin zrzucają liście, łodygi pokryte są korek, który ma wysokie właściwości termoizolacyjne, oraz substancje przeciw zamarzaniu gromadzą się w żywych komórkach.
Wpływ temperatury na organizmy zwierzęce
Temperatura wpływa na zwierzęta poikilotermiczne i homeotermiczne w różny sposób.
Zwierzęta poikilotermiczne są aktywne tylko w okresie optymalnych temperatur dla ich aktywności życiowej. W okresie niskich temperatur zapadają w stan hibernacji (płazy, gady, stawonogi itp.). Niektóre owady zimują jako jaja lub poczwarki. Hibernacja organizmu charakteryzuje się stanem anabiozy, w którym procesy metaboliczne są bardzo silnie zahamowane, a organizm może długi czas iść bez jedzenia. Zwierzęta poikilotermiczne mogą również hibernować pod wpływem wysokich temperatur. Tak więc zwierzęta na niższych szerokościach geograficznych w upalnej porze dnia znajdują się w dziurach, a okres ich aktywnego życia przypada na wczesny poranek lub późny wieczór (lub nocą).
Organizmy zwierzęce zapadają w stan hibernacji nie tylko pod wpływem temperatury, ale także innych czynników. Tak więc niedźwiedź (zwierzę homeotermiczne) zimuje z powodu braku pożywienia.
Zwierzęta homoiotermiczne w mniejszym stopniu zależą od temperatury w swoim życiu, ale temperatura wpływa na nie pod względem obecności (braku) pożywienia. Zwierzęta te mają następujące przystosowania, aby przezwyciężyć skutki niskich temperatur:
1) zwierzęta przemieszczają się z rejonów chłodniejszych do cieplejszych (migracje ptaków, migracje ssaków);
2) zmienić charakter okrywy (letnie futro lub upierzenie zastępowane jest grubszym zimowym; gromadzą się w nich gruba warstwa tłuszczu - dzików, fok itp.);
3) hibernacja (na przykład niedźwiedź).
Zwierzęta homeotermiczne przystosowały się do zmniejszania ekspozycji na temperatury (zarówno wysokie, jak i niskie). Tak więc osoba ma gruczoły potowe, które zmieniają charakter wydzieliny, gdy podwyższonych temperaturach(zwiększa się ilość wydzieliny), zmienia się światło naczyń krwionośnych w skórze (w niskich temperaturach zmniejsza się, w wysokich wzrasta) itp.
Promieniowanie jako czynnik abiotyczny
Zarówno w życiu roślin, jak i zwierząt ogromną rolę odgrywają różne rodzaje promieniowania, które albo wnikają na planetę z zewnątrz (promienie słoneczne), albo są uwalniane z wnętrzności Ziemi. Rozważamy tutaj głównie promieniowanie słoneczne.
Promieniowanie słoneczne jest niejednorodne i składa się z fale elektromagnetyczne różnej długości, a zatem mają różne energie. Powierzchnia Ziemi dociera do promieni zarówno widzialnego, jak i niewidzialnego widma. Widmo niewidzialne obejmuje promienie podczerwone i ultrafioletowe, podczas gdy widmo widzialne ma siedem najbardziej rozróżnialnych promieni (od czerwonego do fioletowego). kwanty promieniowania wzrastają od podczerwieni do ultrafioletu (tj. promienie ultrafioletowe zawierają kwanty najkrótszych fal i najwyższej energii).
Promienie słoneczne pełnią kilka ważnych ekologicznie funkcji:
1) pod wpływem promieni słonecznych na powierzchni Ziemi realizowany jest pewien reżim temperaturowy, który ma charakter strefowy równoleżnikowy i pionowy;
Przy braku wpływu człowieka skład powietrza może się jednak różnić w zależności od wysokości nad poziomem morza (z wysokością spada zawartość tlenu i dwutlenku węgla, gdyż gazy te są cięższe od azotu). Powietrze obszarów przybrzeżnych wzbogacone jest parą wodną, która zawiera sole morskie w stanie rozpuszczonym. Powietrze lasu różni się od powietrza pól zanieczyszczeniami związków wydzielanych przez różne rośliny (na przykład powietrze lasu sosnowego zawiera dużą ilość substancji żywicznych i eterów, które zabijają patogeny, dlatego powietrze to jest lecznicze na gruźlicę pacjentów).
Klimat jest najważniejszym złożonym czynnikiem abiotycznym.
Klimat to skumulowany czynnik abiotyczny, który obejmuje określony skład i poziom promieniowania słonecznego, związany z nim poziom temperatury i wilgotności oraz określony reżim wiatrowy. Klimat zależy również od charakteru roślinności rosnącej na danym terenie oraz ukształtowania terenu.
Na Ziemi istnieje pewna równoleżnikowa i pionowa strefa klimatyczna. Istnieje wilgotny klimat tropikalny, subtropikalny, ostro kontynentalny i inne rodzaje klimatu.
Powtórz informacje o różnych typach klimatu w podręczniku geografii fizycznej. Weź pod uwagę klimat okolicy, w której mieszkasz.
Klimat jako czynnik kumulacyjny tworzy taki lub inny rodzaj roślinności (flory) i blisko spokrewniony typ fauny. Osady ludzkie mają ogromny wpływ na klimat. Klimat dużych miast odbiega od klimatu obszarów podmiejskich.
Porównaj reżim temperaturowy miasta, w którym mieszkasz, i reżim temperaturowy obszaru, w którym znajduje się miasto.
Z reguły temperatura w mieście (zwłaszcza w centrum) jest zawsze wyższa niż w regionie.
Mikroklimat jest ściśle związany z klimatem. Przyczyną powstania mikroklimatu są różnice w rzeźbie terenu na danym terenie, obecność zbiorników wodnych, co prowadzi do zmiany warunków na różne terytoria w tej strefie klimatycznej. Nawet na stosunkowo niewielkim obszarze strefa podmiejska na jego poszczególnych częściach mogą wystąpić różne warunki wzrostu roślin ze względu na różne warunki oświetleniowe.
Nie można przecenić znaczenia wody w życiu roślin. Niezbędne jest utrzymanie strukturalnej integralności komórek, tkanek i całego organizmu. Wszystkie procesy życiowe w roślinach zachodzą w środowisku wodnym. Woda bierze udział w procesie fotosyntezy, w transporcie minerałów. Aby stworzyć 1 część materia organiczna roślina zużywa około 400 części wody. Ilość wody, która dostaje się do roślin, determinuje cały przebieg metabolizmu, zapewnia nie tylko wzrost biomasy, ale także cechy anatomii i morfologii rośliny. Dla glonów woda jest siedliskiem, a dla roślin lądowych ważnym czynnikiem środowiskowym.
W naturze, jak wiadomo, woda występuje w postaci par atmosferycznych, w postaci kropel-ciekłej i stałej krystalicznej. Wilgotność nie jest taka sama w różnych częściach świata. Większość opadów przypada na strefę równikową (od 10° szerokości geograficznej południowej do 10° północna szerokość geograficzna). Zazwyczaj ilość opadów w tej strefie jest różna, ale przeważają obszary z ulewnymi deszczami. Szczególnie dużo opadów pada w górnym biegu Amazonki i na wyspach Archipelagu Malajskiego (Azja Południowo-Wschodnia). Tutaj ilość opadów dochodzi czasami do 12.000 mm rocznie. W Europie najbardziej wilgotnym miejscem są Crkvice nad Adriatykiem (4620 mm rocznie), w Rosji region Batumi (2500 mm rocznie), na Ukrainie - Karpaty (1000 - 1200 mm rocznie). Ale jednocześnie są regiony w tropikach, gdzie ilość opadów jest bardzo mała - nie mogą one nawet zwilżyć powierzchni gleby. Tak więc na pustyni peruwiańskiej i pustyni libijskiej opady wcale nie spadają.
Ważnym warunkiem wpływającym na rozkład opadów jest temperatura. Wraz ze spadkiem temperatury od równika do bieguna maleje intensywność parowania i wilgotność powietrza. W zimnych obszarach powietrze nie jest w stanie pomieścić dużej ilości pary wodnej. W ciepłym natomiast z wody opadającej w postaci deszczu może wykraplać się duża ilość pary. Dużą rolę w redystrybucji opadów na powierzchni ziemi odgrywa obecność dużych zbiorników wodnych. Im bliżej morza lub oceanu, tym więcej opadów spada. Na tę samą redystrybucję wpływa obecność systemów górskich. Powiedzmy na wybrzeżu Pacyfik w Peru i częściowo w Chile jest pustynia Atakama, na której od kilku lat nie pada. Powstał dzięki pasmom górskim, które zatrzymują chmury deszczowe pochodzące z oceanu. Coś podobnego obserwuje się na Ukrainie na Zakarpaciu, gdzie pada nieco więcej niż w regionie Karpat położonym na wschodzie. Oczywiście nie prowadzi to do powstawania pustyń, ale dochodzi do pewnego zatrzymywania wilgoci, co w pewnym stopniu znalazło odzwierciedlenie w tworzeniu zbiorowisk roślinnych położonych na wschód od gór.
Badając ekologię roślin w danym regionie ziemi, należy pamiętać o różnych ilościach opadów w ciągu roku. W strefie równikowej występują dwa okresy opadów marzec - kwiecień i październik - listopad. Ale w Afryce, w dorzeczu Konga, z bardzo dużą ilością rocznych opadów (do 2230 mm), bardzo suchymi okresami roku (lipiec - sierpień), kiedy nie pada deszcz. na średnich szerokościach geograficznych, w tym na Ukrainie, maksymalna ilość Opady występują częściej w lipcu - wrześniu i styczniu - lutym.
Rośliny w różny sposób pobierają wodę w okresie wegetacji. Największa jego ilość jest wchłaniana w obecności wilgotnej gleby, dobrze rozwiniętej powierzchni transpiracyjnej i ciepłego, suchego powietrza. Przykładem są lasy sosnowe porastające wysokie wydmy w klimacie umiarkowanym. W takich warunkach środowiskowych wysoki stopień transpiracji obserwuje się tylko w okresie wegetacji bezpośrednio po deszczu. Po letnich prysznicach wchłaniana jest duża ilość wilgoci, jednak proces ten zatrzymuje się kilka dni po zakończeniu połączonego efektu grawitacji i chłonności korzeni, co skutkuje punkt trwałego więdnięcia.
Wilgotność gleby, przy której rośliny zaczynają wysychać, nazywana jest więdnięciem wilgoci. Zasoby wodne pozostające po punkcie trwałego więdnięcia są trudno dostępne i nazywane są rezerwami martwymi.
W przypadku gleb piaszczystych martwe stado wynosi 1 - 3%, piaszczyste - 5 - 7%, gliniaste - 12 - 15%. Na piaszczystych glebach Polesia, gdzie poziom wód gruntowych wynosi poniżej 3-5 m, sosna zwyczajna obniża pasma systemu korzeniowego w strefę podniesienia kapilarnego i zapewnia mu nawilżenie. Wzrost drzew następuje głównie dokładnie w krótkich okresach. Jednak w większości lasów woda utrzymuje się w glebie przez dłuższy czas. Gromadzi się zarówno z powodu roztopów, jak i opadów deszczu. Ponadto systemy korzeniowe drzew są w stanie dotrzeć do poziomu wód gruntowych. Dlatego w takich warunkach transpiracja, a także wzrost drzew może trwać dłużej.
Cyklony, które tworzą się nad dobrze rozgrzanym oceanem, pobierają duże ilości pary wodnej i przenoszą ją do regionów kontynentalnych, w tym do naszego. Dzieje się tak zimą. Latem aktywność cykloniczna maleje. W regionach kontynentalnych temperatura wzrasta, powstają silne wypory, które również przenoszą wilgoć.
Ocean jest głównym rezerwuarem wody na Ziemi, ponieważ zawiera 91,55% całej wody na Ziemi (tabela 5.1). Organizmy żywe (głównie u producentów, czyli rośliny) zawierają jej znaczną ilość - 1,1 x kg miliardów ton, ale jest to mniej niż 0,0001% jej całkowitej ilości na planecie.
Według M. I. Budyko roczne całkowite parowanie z oceanu wynosi 448 tys. km3 wody, a z lądu, zwłaszcza w wyniku transpiracji roślinności - 71 tys. W wyniku opadów do lądu dostaje się 109 tys. km3 wody, a do oceanu 411 tys. km3. Nawiasem mówiąc, z sumy opadów na lądzie 37 tys. km3 wraca do oceanu. Tak więc w przyrodzie istnieje stały obieg wody, który wraz z innymi czynnikami określa klimat terytoriów, a także stwarza warunki do życia i żywotnej aktywności roślin (ryc. 5.1).
Tabela 5.1
Masa wody w hydrosferze i jej składnikach
|
składniki hydrosfery |
Masa wody (10 3 miliardy ton) |
Udostępnij z (%): |
czas kompletny powrót do zdrowia (lata) |
Konwencjonalnie warstwa na powierzchni Ziemi (m) |
|
|
rezerwy wody słodkiej |
ogólny szerokie rzesze |
||||
|
świat Ocean |
|||||
|
Wody gruntowe |
|||||
|
w tym świeże |
|||||
|
Formacje śnieżne i lodowe |
|||||
|
Małe elementy: |
|||||
|
jeziora |
(świeży) |
(świeży) |
|||
|
wilgotność gleby |
|||||
|
bagna |
|||||
|
wilgotność powietrza |
|||||
|
rzeki |
|||||
|
biologiczny |
|||||
|
CAŁKOWITY |
1500496,3 |
||||
Ryż. 5.1. Obieg wody w biogeocenozach lądowych
(Według A. A. Molchanova, 1964)
Parowanie z powierzchni ziemi należy rozpatrywać w połączeniu z opadami atmosferycznymi. Dla bilansu wodnego dowolnego terytorium ważne jest nie tylko Wartości bezwzględne opady i parowanie, jaki jest stosunek między nimi.
Jeżeli ilość rocznych opadów przekracza ilość parowania w tym samym okresie, wówczas terytorium to należy do strefy wilgotnej, w przeciwnym razie - do strefy suchej.
Około 1/3 powierzchni lądu cierpi na niedobory wody, podczas gdy na połowie tego obszaru panuje skrajnie sucha sytuacja z rocznymi opadami poniżej 250 mm i parowaniem powyżej 1000 mm. Niewielka ilość opadów sama w sobie nie wskazuje jeszcze na suchość terytorium: zimne strefy polarne są również ubogie w opady, ale nie są suche, ponieważ parowanie jest tutaj bardzo niskie. Regiony wyjątkowo wilgotne wysokie poziomy opady zajmują mniej niż 9% powierzchni ziemi. Znaczące obszary suche znajdują się głównie między 15° a 30° szerokości geograficznej północnej i południowej oraz za wysokimi pasmami górskimi. Wraz z oddaleniem od morza następuje stopniowe przejście ze strefy wilgotnej do suchej (ryc. 5.2). Ten ostatni charakteryzuje się suszą i zasoleniem gleb i wód. Jednak stosunek rocznych opadów i parowania jest raczej przybliżonym wskaźnikiem wilgotności lub suchości terytorium. W przypadku roślin ważne jest, aby wilgoć była dostarczana dokładnie wtedy, kiedy są najbardziej potrzebne, czyli w okresie wegetacji. Dla bardziej obiektywnego wniosku, zgodnie z oceną klimatu na danym obszarze, konieczne jest sporządzenie wykresów klimatycznych w oparciu o obserwacje meteorologiczne w ciągu roku.

Ryż. 5.2. Schemat zmian klimatu, roślinności i gleb wzdłuż profilu przez główne krajobrazy europejskiej części Rosy z północnego zachodu na południowy wschód
(Według G. M. Wysockiego i G. F. Morozowa, 1934)
Uwaga: horyzont próchniczy zaznaczono na czarno, horyzont iluwium zaznaczono paskami.
Przykładem przewagi parowania wilgoci nad ilością opadów mogą być pustynie Azji Środkowej, które obejmują część Kazachstanu, południową część Niziny Aralsko-Kaspijskiej i rozciągają się na wysokie góry na wschodzie. Na południu wkraczają do Afganistanu. Opady w południowej części pustyń Azji Środkowej padają głównie zimą, a latem w ogóle się nie zdarzają. Roczna ilość opadów nie przekracza 250 mm, ale na dużych obszarach zmniejsza się do 100 mm lub mniej. Niskie parowanie wiąże się z niskimi temperaturami zimą: 1000 - 1400 (do 2500) mm, dzięki czemu roślinność rozwija się na piaszczystych pustyniach, nawet przy bardzo głębokich wodach gruntowych. Na pustyniach Azji Środkowej niektóre formacje roślinne są związane z ilością opadów i charakterystyką ich rozmieszczenia w ciągu roku, a także z podłożami glebotwórczymi:
Efemeryczna pustynia na lessowych, gliniastych glebach;
Zgrupowania roślin kamienistej pustyni gipsowej
Formacje halofitów na glebach gliniastych;
Pustynia Psamophyte na glebach piaszczystych.
I tak na przykład efemeryczne pustynie zajmują rozległe obszary na zachód od jeziora Bałchasz (Głodny Step) (tab. 5.2). W marcu i kwietniu w regionie pada co 4-5 dni, dobrze nawilżając górne poziomy gleby. Pod koniec maja gleba wysycha i nagrzewa się do głębokości 10 cm do + 30 ° C i więcej. Wiosną szybko rozwijają się tu gatunki roślin, które tworzą podziemne byliny (maczugowce, cebulki, kłącza). To jest turzyca gospodarza (Carex hostii), bulwiasty bluegrass ( Poa bulbosa L.), bulwiasta geranium (Geranium tuberosum L.), a także rodzaje rodzajów lub zmyachka (Skorzonera) gwiazdki (Gagea) tulipan (Tilira) i inne Wiosenna pustynia przypomina kwitnącą łąkę – szata roślinna jest całkowicie zamknięta, a rośliny nie mają cech kseromorficznych. W niektórych miejscach można zobaczyć gigantyczną roślinę zielną z rodziny parasolowatych - śmierdzącą ferulę (Ferula foetida), który osiąga wysokość do 2 m. Kwiaty na tej roślinie pojawiają się 6-7 lat po wykiełkowaniu nasion. Po owocowaniu umiera. Ale właśnie w cechach korzenia feruli przejawia się adaptacja tego gatunku do suchych warunków. Roślina tworzy potężny macicaŚrednica 17 - 29 cm, która wnika na głębokość zaledwie 20 cm, a ponadto dzieli się na liczne korzenie boczne, które z kolei rosną poziomo do 2 m długości, a następnie pogłębiają się w pionie o 80 - 100 cm Tak więc korzenie jednej rośliny penetrują do 4 m3 gleby. Taka struktura korzeni zapewnia roślinie możliwość pozyskiwania wody z powierzchni i nawilżenia głębokich poziomów gleby, a potężny korzeń palowy pomaga przetrwać bezwodne okresy w roku. Innym przykładem adaptacji do suchego klimatu jest bulbastia bluegrass, której cebulki tracą zdolność kiełkowania nawet po 8 latach przechowywania w stanie suchym. Ale od końca kwietnia przez kolejne 9 miesięcy pustynia wygląda na całkowicie wymarłą. Wyjątkiem są dwa małe gatunki zielne Afanopleura pilar (Aphonopleura capillifolia)) i pęcherzyków dwumianowych (Diarthron vesiculosum). Ich kwiaty pojawiają się, gdy słońce wypala wszystkie żywe istoty, a gleba bez wody staje się twarda jak kamień. Jednocześnie pęcherz dwumianowy wysycha dopiero jesienią (to zjawisko ekologiczne nie jest dobrze poznane).
Tabela 5.2
Dynamika zawartości wody w glebie Głodnego Stepu wiosną (wilgotność, %)
Typowy przykład pustyń środkowoazjatyckich wyraźnie pokazuje, że większość fitomas ich roślin jest skoncentrowana w glebie. To wystarczy duże drzewa. W efemerach system korzeniowy jest słaby i zlokalizowany w najwyższa warstwa gleba. Inne gatunki mają zarówno korzenie rozłożone poziomo, jak i te, które wnikają głęboko i docierają do niższych poziomów gleby, zawierając wilgoć dostępną dla roślin nawet latem. Gwarantuje to tym gatunkom pewien zapas wody przez cały rok. Szybkość transpiracji rośliny pustynne zależy od ilości wchłoniętej wody. Przy wystarczającej ilości wilgoci rośliny energicznie transpirują, co samo w sobie jest oznaką intensywnej wymiany gazowej oraz intensywnej fotosyntezy i produkcji materii organicznej. Najwięcej wilgoci na transpirację rośliny wydają w lipcu. W okresie letnim, wraz z pogorszeniem warunków zaopatrzenia w wodę i wzrostem niedoboru wody, tempo parowania maleje. Przejawia się to tym, że wszystkie gatunki zmniejszają obszar transpiracji poprzez zrzucanie części liścia.
Za pomocą stopień i charakter dostarczania wilgoci Rośliny pustynne dzielą się na cztery grupy:
hydrofity, związane z wodami gruntowymi, które są stosunkowo wysoko nad powierzchnią lub nawet wypływają na powierzchnię;
freatofity, które rozwijają się w mikrostacji z niższą lokalizacją wód gruntowych;
trichohydrofity, którego system korzeniowy nie sięga poziomu wód gruntowych, ale wnika w krawędź kapilarną;
ombrofity, które żywią się wyłącznie wodą deszczową.
Tak więc w reżimie wodnym roślin jest bardzo znaczenie odgrywa wilgotność gleby. To tutaj rośliny otrzymują najwięcej wody.
Ilość wilgoci w glebie, jej dostępność determinują przebieg wielu procesów fizjologicznych roślin, rytm ich rozwoju oraz wpływają na ich wygląd.
Zużycie wody przez rośliny z gleby zależy od chłonności ich systemu korzeniowego, od stanu nawilżenia gleby, od sił zatrzymujących wodę w glebie. Cała wilgotność gleby dzieli się na dostępne i niedostępne dla roślin:
Dostępne obejmuje grawitacyjne (wypełnia przerwy między cząsteczkami gleby) i kapilarne (zawarte w kapilarach glebowych). Ten ostatni dostarcza najwięcej wody roślinom;
Niedostępna wilgotność gleby obejmuje: film (adsorpcję), który pokrywa cząstki gleby, higroskopijny i koloidalny.
Różne gleby w różny sposób zatrzymują wilgoć. mają najwyższą wilgotność gleby gliniaste, najmniejszy - piaszczysty (tab. 5.3). Szybkość wznoszenia kapilarnego jest odwrotnie proporcjonalna do wielkości cząstek: im są one mniejsze, tym większa wysokość wznoszenia kapilarnego, ale tym wolniej. Dostępność wilgoci w glebie jest ograniczona kilkoma czynnikami:
Niska temperatura gleby;
Wysokie stężenie rozpuszczalnych soli;
Wysoka kwasowość gleby;
Brak powietrza glebowego;
Brak tlenu w wodach gruntowych.
Tabela 5.3
Zależność wysokości podciągu wody od składu mechanicznego gleby
(Według A. A. Rode, 1963)
|
Wielkość cząstek gleby, mm |
Wysokość podnoszenia wody, mm |
Badając ekologiczną rolę wody, należy zawsze pamiętać o równowadze między poborem a zużyciem wody. Przyjęcie często następuje przez system korzeniowy, a uszkodzenie - z powodu transpiracji. Jeśli dopływ wilgoci nie rekompensuje kosztów, roślina umiera. Dzieje się tak, gdy w glebie brakuje wilgoci, choroby roślin, wzrost temperatury, suche wiatry. W zależności od czasu działania jednego lub drugiego negatywnego czynnika w roślinach zachodzą różne zmiany, które prowadzą najpierw do spowolnienia ich rozwoju, a następnie do śmierci:
Zmniejsza się turgor komórek:
Zatrzymuje podział i wzrost komórek; aparaty szparkowe zamykają się i dopływ CO2 do roślin ustaje;
Zmniejsza się intensywność fotosyntezy;
Intensywność oddychania wzrasta;
Materia organiczna jest zużywana;
Transpiracja jest bardzo powolna;
Wypływ składników odżywczych z gleby spowalnia i całkowicie się zatrzymuje;
Następuje śmierć ssących korzeni i tym podobnych.
W różne okresy rośliny na różne sposoby tolerują brak wilgoci, taki okres nazywa się krytycznym. Większość roślin jest najbardziej podatna na powstawanie organów generatywnych. Różne rośliny w tych samych warunkach inaczej tolerują brak wilgoci w glebie. Zależy to od siły ssącej komórek systemu korzeniowego. Na przykład rośliny tundry rosnące na glebach zasolonych iw regionach suchych wyróżniają się wysokim ciśnieniem osmotycznym komórek korzeniowych. Jeden i ten sam gatunek, który rośnie w tundrze i na średnich szerokościach geograficznych, ma siłę ssącą o 10-20% większą na północy. Rozmiar systemu korzeniowego ma znaczenie. Tak więc roczne sadzonki dębu mają całkowitą długość korzenia 114 m, jesion - 186 m, natomiast długość części nadziemnych wynosi 20 - 25 cm U niektórych roślin pszenicy w okresie dojrzewania nasion łączna długość korzenie osiągają 64 - 80 km. Ważna jest również głębokość penetracji korzeni w glebie. Na przykład długość korzeni cierniowych wielbłądów (ryc. 5.3) poza lucerną może sięgać warstw wodonośnych na głębokości 18 m. Po osiągnięciu horyzontu system korzeniowy rozwija się na głębokości.

Ryż. 5.3. System korzeniowy wielbłąda ciernie i sosna zwyczajna (a - bagno torfowiec, b - na glebie piaszczystej)
Kolejnym czynnikiem determinującym bilans wodny roślin jest jej wydatek zachodzący podczas transpiracji. Rośliny wykształciły adaptacje ochronne przed wysokimi temperaturami i brakiem wody:
Zdolność do zamykania aparatów szparkowych przy braku wody lub ich głębokiej lokalizacji w liściach;
Opadające liście i pojedyncze pędy (dzhuzgan, saxaul)
Zmniejszenie obszaru transpiracji (w piórach liście trawy są skręcone w rurkę)
Częściowa lub całkowita redukcja liści (ciernie kaktusów i cierń wielbłąda, szydłowaty wyrastają w saxaul biały, łuski u jałowca kozackiego)
Pojawienie się adaptacji ograniczających parowanie lub zatrzymujących wilgoć transpiracyjną (grube włoski na liściach dziewanny, błyszcząca powierzchnia liści borówki brusznicy, błyszczące łuski na srebrzystej przyssawce, parowanie olejek eteryczny w celu zmniejszenia nagrzewania się blachy, kryształów soli mineralnych w solance, tworzenia się wosku i powłoki żywicznej w celu zmniejszenia parowania, tworzenia zwartych tkanek powłokowych)
Regulacja parowania przez dobowe ruchy pędów i liści (dziki molokan)
Restrukturyzacja rytmów życia (efemerydy)
Okresowe wstrzymywanie i wznawianie rozwoju kilkukrotnie w okresie wegetacji (niektóre gatunki turzyc)
Zdolność do gromadzenia wilgoci w ich narządach wegetatywnych (aloes, kaktusy).
Tak więc w procesie ewolucji rośliny rozwinęły różne adaptacje do warunków istnienia związanych z wilgocią. Dlatego zaczęto je dzielić na pewne grupy wymagań dotyczących wilgotności.
Hydrofity (z gr. Higros – woda, fiton – roślina) – rośliny rosnące w środowisku wodnym.
Są to rośliny, które rosną częściowo lub całkowicie zanurzone w wodzie, takie jak rzęsa, ożypałka, grot strzały, lilie wodne, dzbany. Ponieważ w wodzie powstaje określony reżim temperatury, reżim oświetlenia i nasycenia gazami i składnikami odżywczymi rozwinęły się hydrofity konkretne oprawy:
Rozwinięta stosunkowo duża powierzchnia liści;
Wszystkie narządy pokryte śluzem, aby zapobiec wypłukiwaniu soli;
Słabo rozwinięte tkanki mechaniczne;
Wszystkie narządy mają objętość powietrza;
Słabo rozwinięte tkanki powłokowe ułatwiają wchłanianie gazów i składników odżywczych z wody;
Liście zanurzone w wodzie mają typową strukturę cienia, a te znajdujące się na powierzchni mają strukturę jasną;
Słaby rozwój systemu korzeniowego, aż do całkowitego zaniku;
Rozmnażanie wegetatywne jest wysoko rozwinięte.
Hygrophytes - (z greckiego Hidros - mokry, fiton - roślina) - Rośliny lądowe które rosną w warunkach wysoka wilgotność(gleba lub powietrze).
Mają głównie cienkie, duże blaszki liściowe z słabo rozwiniętym naskórkiem, słabo rozwiniętym systemem korzeniowym i cienkimi łodygami. Rosną na terenach podmokłych, w wilgotnych lasach, na terenach zalewowych, wzdłuż brzegów rzek i zbiorników wodnych. Wśród znanych nam roślin są to olsza czarna, żurawina błotna, zegarek trójlistny (zegarek), nagietek błotny, fiołek błotny, turzyca, wierzba bagienna. Ponieważ nie regulują one dobrze transpiracji, bardzo trudno jest wytrzymać suszę. Rekultywacja drenażowa prowadzi do ich śmierci.
Mesofity (z greckiego Mesos - średni, pośredni, fiton - roślina) - ekologiczna grupa roślin, która rośnie w warunkach średniej wilgotności.
Różnią się one różnorodnymi formami i nie mają tak specyficznych cech morfologicznych i anatomicznych jak kserofity i higrofity. Do tej grupy można zaliczyć wiele roślin: świerk, buk, borówka brusznica, golteria, kopytko, tymotka, kostrzewa, konwalia, osika, brzoza, lipa, klon, owies, ziemniaki, jabłonie itp. Rośliny te dominują w umiarkowane warunki klimatyczne.
Kserofity (z gr. Xeros - suchy, pośredni, fiton - roślina) - rośliny o suchych miejscach, przystosowane do życia w warunkach długotrwałej lub sezonowej gleby i suszy atmosferycznej.
Posiadają specjalne urządzenia, które zapobiegają parowaniu wody i zapobiegają przegrzewaniu się roślin. Przez długi czas dominowała teoria niemieckiego naukowca A. Schimpera, że kserofity wykorzystują wodę ekonomicznie, czyli charakteryzują się niską intensywnością transpiracji. Ale w 1952 roku radziecki naukowiec N.A. Maksimov udowodnił, że główną różnicą między mezofitami a kserofitymi nie jest intensywność transpiracji, ale zdolność roślin odpornych na suszę do wytrzymania odwodnienia podczas suszy. Rosną na stepach i pustyniach. Do znanych gatunków tej grupy roślin należą puszysty dąb, sosna krymska, trawa piórkowa, piołun, bluegrass. Na północy Ukrainy w górnych partiach wydm występują kserofity (szałwia, różne gatunki piołunu). Kserofity dzielą się na dwie grupy:
sukulenty(z greckiego Succulentus - soczysty)
sklerofity(z greckiego Skleros - twardy, twardy).
Niektóre cechy sukulentów zostały omówione podczas scharakteryzowania temperatury jako czynnika środowiskowego. W przypadku niektórych sukulentów liście zamieniły się w kolce lub łuski. Funkcje asymilacyjne pełnią zielone łodygi. Przeciwnie, w formach liściowych nie ma łodyg. Pędy i liście sukulentów są często nagie, pokryte grubą skórką i naskórkiem woskowym. Ich korzenie szybko rosną, osiągają znaczne rozmiary i występują powszechnie w powierzchniowej warstwie gleby. Moc absorpcji korzeni w sukulentach jest niska. Rośliny z tej grupy charakteryzują się specyficznym metabolizmem. W procesie oddychania cukry nie rozkładają się na CO2 i H2O, jak w większości roślin, ale kumulują się substancje pośrednie - kwasy organiczne. W ciągu dnia kwasy te rozkładają się, uwalniając dwutlenek węgla, który jest natychmiast wchłaniany przez samą roślinę. Tkanki asymilacyjne reprezentowane są głównie przez miąższ kanalikowy, pośrodku którego znajduje się miąższ zatrzymujący wodę.
Do sklerofitów zalicza się saksaul, różne rodzaje traw i piołunów, cierń wielbłąda. Ze względu na dużą ilość tkanek mechanicznych rośliny te są twarde, mogą stracić do 25% swojej wilgoci. System korzeniowy ma dużą siłę wchłaniania. W strukturze jest inny, ale częściej wchodzi bardzo głęboko w warstwy wodonośne gleby. Typowy sklerofit to skarłowaciała roślina o długich korzeniach. Na przykład cierń wielbłąda ma wysokość do 1 m, a jego system korzeniowy ma długość do 30 m. Liście są albo zmniejszone, albo małe, pokryte gęstą skórką, często owłosione i mogą się zwijać. Uprawa kserofitów w ciepłym, ale wilgotnym klimacie prowadzi do większej intensyfikacji wzrostu, wzrostu wielkości liści. Zamiast kolców może się rozwijać miękkie liście. Wskazuje to, że rośliny te zostały wyparte przez inne gatunki w niesprzyjających warunkach wzrostu.
W obrębie tej samej strefy klimatyczno-przyrodniczej oraz w obrębie określonych fitocenoz rośliny są również podzielone na grupy według ich wymagań wilgotnościowych. Przykładem jest skład gatunkowy fitocenoz leśnych w strefie stepowej Ukrainy. Składa się ona z:
Kserofity - sosna zwyczajna, Gleditsia trokhkolyuchkova, robinia akacjowa, przyssawka, dąb puszysty, sosna krymska, ailanthus;
Mezokserofity – kora brzozy (forma korkowa), dzika róża, migdał, tarnina;
Xeromesophytes - dąb szypułkowy, wilcza jagoda, jesion wyniosły, gruszka, jabłoń;
Mezofity - grab, leszczyna, wiąz, lipa pospolita, klon zwyczajny, modrzew syberyjski, jawor;
Mesohygrofity - topola biała i czarna, osika, brzoza omszona, kruszyna łamliwa, kalina, czarny bez;
Hygrofity - wierzba biała, wierzba krucha, olcha czarna, czeremcha.
Ciekawe cechy ekologiczne mają wspólne gatunki drzew w lasach Polesia Ukrainy - sosna zwyczajna (Pinus sylvestris). Z powodzeniem rośnie w różnych (pod względem wilgotności gleby i głębokości wód gruntowych) warunkach środowiskowych – od wierzchołków wydm na równinach sandrowych po bagna oligotroficzne i mezotroficzne (ryc. 5.4). Oznacza to, że zarówno tam, gdzie poziom wód gruntowych leży na głębokości 4-5 m lub więcej, jak i tam, gdzie dochodzi do powierzchni. Zazwyczaj sosna zwyczajna jest określana jako kserofity, co tłumaczy się jej zdolnością do dość dobrej tolerancji znacznego suchego powietrza i wzrostu na suchych glebach. Na przykład to drewno z powodzeniem tworzy nasadzenia na stepie Ukrainy. Ale ponieważ rośnie nawet w najbardziej ubogich w składniki odżywcze warunkach glebowych (lasy sosnowe) i w różnych reżimach wilgotnościowych, można go przypisać prawie każdej z powyższych grup roślin. Sosna tworzy jednak najbardziej wydajne, zamknięte plantacje, gdy poziom gruntu znajduje się 1,5–2 m bliżej powierzchni gleby niż w warunkach, w których rosną typowe kserofity.

Ryż. 5.4. Zależność wzrostu sosny na glebach piaszczystych od głębokości wód gruntowych
(Według P. S. Pogrebniaka)
A1 - las sosnowy porostowy (Pinetum cladinosum) porośnięty porostami
A2 - zielony mchowy las sosnowy ( Pinetum hylocomiosum) z żurawiną (Vaccinium vitis-idaea)
A3 - borówkowy las sosnowy (Pinetum myrtillosum) z domieszką brzozy o głębokości wód gruntowych 1 - 2 m A4 - bór buchowo-bagnowy (Pinetum uligino-ledosum) na glebach torfowo-glejowo-bielicowych A5 - torfowiska wysokie z rzadkimi sosnami na torfowiskach "pijanych"
Rośliny dostosowują się nie tylko do ogólnej wilgotności terenu, ale także do wahań sezonowych i krótszych. Dostosowując się do krótkotrwałych wahań warunków zaopatrzenia w wodę i parowania, G. Walter podzielił rośliny lądowe na dwie grupy:
Poikilohydric, o zmiennej zawartości wody w tkankach;
Homojohydrat. które są w stanie utrzymać względnie stałą zawartość wody w tkankach.
Pierwsza grupa obejmuje bakterie, sinice, porosty, niektóre rodzaje mchów i zarodników roślin naczyniowych, niewielką liczbę okrytozalążkowych. Są to prokarionty lub eukarionty, które mają małe komórki bez centralnej wakuoli. Po wysuszeniu kurczą się bez zmiany ultrastruktur protoplazmy, a tym samym bez utraty żywotności. Wraz z utratą wody osłabiają się różne funkcje życiowe, takie jak fotosynteza i oddychanie. Rośliny te wchłaniają wodę z wilgotnego podłoża lub po zwilżeniu deszczem, rosą i mgłą w sposób kapilarny i pęcznieją. Kiedy roślina pęcznieje, przywracane są w niej normalne procesy metaboliczne.
Druga grupa obejmuje większość roślin. ich komórki mają dużą centralną wakuolę, a dzięki obecności wewnętrznego środowiska wodnego protoplazma staje się mniej zależna od warunki zewnętrzne. Ale taka komórka, a zatem i roślina jako całość, straciła zdolność wysychania. Rośliny przypisane do grupy muszą rosnąć w miejscach o mniej lub bardziej stałym źródle wilgoci.
Omawiając wilgotność jako czynnik środowiskowy, należy zwrócić uwagę na fakt, że nie tylko wilgoć wpływa na rośliny, ale rośliny w dużej mierze determinują wilgotność fitocenozy. Od dawna wiadomo, że lasy wpływają na wilgotność powietrza w fitocenozie i na terenach przyległych, wilgotność gleby oraz poziom wód gruntowych.
Podczas deszczu wilgoć utrzymuje się na liściach, pędach, pniach drzew. Przy lekkim deszczu korona jodły zatrzymuje 80% opadów, a sosna - 60 %. Drzewa liściaste zachowują znacznie mniej wilgoci z deszczu. Nawet w gęstym bukowym lesie około 50% wody deszczowej dociera do gleby. Dużo wilgoci zatrzymuje się również w fitocenozach ziołowych. Na przykład na łąkach regionu Petersburga tylko jedna trzecia opadów dociera do gleby. Część wilgoci trafia do zwilżenia organów wegetatywnych roślin, a część koncentruje się na ich powierzchni w postaci kropli.
Wilgotność powietrza w fitocenozach leśnych jest nieco wyższa niż na sąsiednich terenach ze względu na znaczną transpirację wilgoci przez rośliny, zmniejszenie siły wiatru, spadek wysokich temperatur i zmniejszenie parowania z powierzchni gleby. Latem, na gęstych plantacjach liściastych, wilgotność przy powierzchni gleby jest wyższa niż w rejonie koszenia o prawie 15–20% (Molchanov, 1961). W zbiorowiskach zielnych zjawisko to obserwuje się tylko częściowo, w niewielkiej warstwie powietrza przy powierzchni gleby. Wilgotność jest bardzo zróżnicowana w zależności od rodzaju lasu. Tak więc latem w czystych lasach sosnowych piaszczysta gleba jest o 10 - 12% wyższy niż w lasach dębu roboczego. W lesie wilgotność względna powietrze maleje wraz z wysokością, w koronach osiąga pewną stałą wartość, a najmniejsze - przy wychodzeniu z koron. Specyficzne warunki wilgotności powietrza (wraz z innymi warunkami środowiskowymi) w fitocenozach leśnych lasów tropikalnych wyjaśniają obecność w nich epifitów.
Mają przystosowania do wydobywania składników odżywczych i wody ze środowiska (np. gąbczaste okrywy korzeniowe, które łatwo wchłaniają wodę, Specjalna edukacja z liści i pędów do zatrzymywania wody itp.).
Fitocenozy wydzielają bardzo dużą ilość wilgoci. Las w pewnych warunkach może wyparować więcej wody niż w ciągu roku. Na przykład dojrzały las bukowy sezon wegetacyjny do atmosfery przedostaje się od 3600 do 5400 ton wody z 1 ha. Jest to rodzaj pompy, której działanie wpływa na reżim hydrologiczny nie tylko terytorium, na którym rośnie, ale także terytoriów sąsiednich. Na przykład liczni przedstawiciele drzew eukaliptusowych wyróżniają się zdolnością do intensywnego wchłaniania i odparowywania wilgoci. Nazywa się je nawet „pumpami”. Szacuje się, że w ciągu roku jedno drzewo eukaliptusowe może odparować do 14 ton wody. Ta umiejętność służy do osuszania terenów podmokłych. Najbardziej znanym przykładem jest sadzenie drzew eukaliptusowych w celu osuszenia Niziny Colchis (kaukaskie wybrzeże Morza Czarnego) podczas walki z malarią w ZSRR.
Na sucho strefy klimatyczne Plantacje leśne służą do zatrzymywania śniegu, przenoszenia spływu powierzchniowego do spływu podpowierzchniowego, zwiększając w ten sposób wilgotność gleby.
Na Ukrainie występuje duża liczba bagien i terenów podmokłych (zwłaszcza na Polesiu), na których tworzą się specyficzne fitocenozy i rosną różne rośliny higrofitowe.
Bagno to obszar powierzchni ziemi charakteryzujący się nadmierną wilgocią, nagromadzeniem nierozłożonej materii organicznej i obecnością roślinności kochającej wilgoć.
Bagna powstają podczas zarastania zbiorników wodnych, zasypywania lasów i pożarów lasów, cebuli i tym podobnych. W zależności od warunków ulgi występowania i odżywiania wodno-mineralnego bagna dzielą się na:
nisko położony (eutroficzny)
przejściowy (mezotrony)
jazda konna (oligotroficzna).
Bagna nizinne (eutroficzne) charakteryzuje się najbogatszymi glebami. Szata roślinna jest tu bogata i urozmaicona i składa się z brzozy omszonej, olchy lepkiej (czarnej), wierzby krzewiastej, trzciny, kugi, skrzypów, turzyc, zegarków, pięciorników, mchów hypnum itp. Podniesione (oligotroficzne) bagna ubogie w składniki mineralne, rozwijają się głównie w strefie leśnej. Z higrofitów tutaj sosna pospolita, brzoza omszona, niektóre rodzaje turzycy, wełnianka pochwowa, żurawina, mchy torfowce. Te ostatnie są podstawą torfowisk wysokich. Mchy torfowce są w stanie wchłonąć wodę. Około połowa komórek torfowca jest pusta. To oni są wypełnieni wodą, zwiększając masę rośliny od 20 do 50 razy. Łodyga tego mchu rośnie w górę, osiągając długość ponad 150 cm Ciągły wzrost i wysoka pojemność wodna torfowca jest ważnym czynnikiem ekologicznym. Zmusza rośliny kwitnące torfowisk wysokich do przystosowania się do wzrostu torfowiska na wysokość. Mogą tu rosnąć tylko te rośliny, których kłącza mogą również rosnąć na wysokość lub takie, które mogą tworzyć dodatkowe korzenie na pędach (ryc. 5.5). Jednak fitocenozy wytworzone na torfowiskach wysokich charakteryzują się niską biomasą i małą różnorodnością roślin. Rośliny, w tym drzewiaste, są niewymiarowe, a gęstość sadzenia jest poniżej średniej. Bagna przejściowe (mezotroficzne) charakteryzują się umiarkowanym odżywianiem mineralnym i zajmują pozycja pośrednia między niskim a wysokim. Szatę roślinną charakteryzuje brzoza omszona, sosna, niektóre turzyce, błoto, żurawina, wełnianka pochwowa i mchy torfowce.

Ryż. 5.5. Cechy strukturalne roślin torfowiskowych wysokich
(Po Beret, 1927)
a - rosiczka okrągłolistna z pokrywy torfowca, poziom położenia jej rozety wzrasta o 3 cm rocznie
b - świerk w wieku 9 lat, którego pień zakopany jest pod mchem na głębokość 20 cm
Rozkład fitocenoz na Globus z zastrzeżeniem pewnych wzorców, które zależą od stosunku czynników środowiskowych. Dość pełny obraz zapewnienia wegetacji na pewnym terytorium wilgocią podaje różne współczynniki, biorąc pod uwagę kilka wskaźników klimatycznych, w tym opady lub wilgotność. Obejmują one współczynnik hydrotermalny G. T. Selyaninova (GTK), co definiuje się jako stosunek ilości opadów w okresie wegetacji do ilości aktywnych temperatur (powyżej +10 ° C) w tym samym czasie:
gdzie R jest ilością opadów w okresie, kody t °> 10 ° С;
Σ (t ° > 10 ° С) - suma aktywnych temperatur powyżej 10 ° С.
Oznacza to, że przy obliczaniu współczynnika hydrotermalnego Selyaninova brane są pod uwagę dwie cechy warunków środowiskowych - ilość opadów i temperatura powietrza. Powiedzmy, że północna granica stepu pokrywa się z wartością HTC = 1, dla pustyni HTC = 0,5, dla lasów liściastych HTC = 1,5. Zatem dla strefy nadmiernej wilgoci HTC > 1,5, gdy HTC waha się w zakresie od 1 do 1,5, tworzone są dostateczne warunki do nawilżania; przy wartościach GKT od 0,7 do 1 brakuje wilgoci.
Podsumowując charakterystykę wilgotności jako czynnika środowiskowego, należy zauważyć, że w prawie tych samych warunkach geograficznych na Ziemi występują zarówno suche pustynie, jak i lasy deszczowe(rys. 5.6). Różnice przejawiają się w ilości opadów, które spadają na terytorium w ciągu roku od 0,2 - 200 mm na pustyni do 900 - 2000 mm w tropikach. W pierwszym przypadku szata roślinna charakteryzuje się niewielką liczbą gatunków i ich niską produktywnością, w drugim przypadku fitocenozy są złożone w budowie, liczne pod względem różnorodności gatunkowej i ogromne w biomasie.

Ryż. 5.6. Zależność typów roślinności od warunków klimatycznych
(Według M. I. Nikolaykina, N. E. Nikolaykina, A. P. Melekhov, 2004)
Zatem to właśnie wilgotność jest jednym z czynników środowiskowych determinujących rodzaj zgrupowania roślin. W procesie ewolucji rośliny wypracowały szereg specyficznych adaptacji mających na celu zarówno maksymalne wykorzystanie wody, jak i jej nadmiar lub brak. Ponadto środowisko wodne jest siedliskiem dużej i ważnej grupy roślin dolnych glonów.
Wstęp
1. Światło jako czynnik środowiskowy. Rola światła w życiu organizmów
2. Temperatura jako czynnik środowiskowy
3. Wilgotność jako czynnik środowiskowy
4. Czynniki edaficzne
5. Różne środowiska życia
Wniosek
Lista wykorzystanej literatury
Wstęp
Na Ziemi panuje ogromna różnorodność warunków środowiska życia, co zapewnia różnorodność nisz ekologicznych i ich „zasiedlenie”. Jednak pomimo tej różnorodności istnieją cztery jakościowo różne środowiska życia, które mają określony zestaw czynników środowiskowych i dlatego wymagają określonego zestawu. adaptacje. Są to środowiska życia: ziemia-powietrze (ląd); woda; gleba; inne organizmy.
Każdy gatunek jest przystosowany do określonego zestawu warunków środowiskowych - niszy ekologicznej.
Każdy gatunek jest przystosowany do swojego specyficznego środowiska, do określonego pokarmu, drapieżników, temperatury, zasolenia wody i innych elementów. świat zewnętrzny bez którego nie może istnieć.
Do istnienia organizmów wymagany jest kompleks czynników. Zapotrzebowanie organizmu na nie jest inne, ale każde w pewnym stopniu ogranicza jego istnienie.
Brak (brak) niektórych czynników środowiskowych może być skompensowany przez inne bliskie (podobne) czynniki. Organizmy nie są „niewolnikami” warunków środowiskowych – do pewnego stopnia same dostosowują się i zmieniają warunki środowiskowe w taki sposób, aby złagodzić brak pewnych czynników.
Braku fizjologicznie niezbędnych czynników (światła, wody, dwutlenku węgla, składników odżywczych) w środowisku nie można zrekompensować (zastąpić) innymi.
1. Światło jako czynnik środowiskowy. Rola światła w życiu organizmów
Światło jest jedną z form energii. Zgodnie z pierwszą zasadą termodynamiki, czyli zasadą zachowania energii, energia może zmieniać się z jednej postaci w drugą. Zgodnie z tym prawem organizmy są systemem termodynamicznym, który nieustannie wymienia energię i materię ze środowiskiem. Organizmy na powierzchni Ziemi narażone są głównie na przepływ energii energia słoneczna, a także długie fale promieniowanie cieplne ciała kosmiczne. Oba te czynniki determinują warunki klimatyczneśrodowisko (temperatura, szybkość parowania wody, ruch powietrza i wody). Na biosferę pada z kosmosu światło słoneczne o energii 2 cal. na 1 cm2 w 1 min. To tak zwana stała słoneczna. Światło to, przechodząc przez atmosferę, jest osłabione i nie więcej niż 67% jego energii może dotrzeć do powierzchni Ziemi w pogodne południe, tj. 1,34 cal. na cm2 w 1 min. Przechodząc przez zachmurzenie, wodę i roślinność światło słoneczne ulega dalszemu osłabieniu, a rozkład energii w nim w różnych częściach widma zmienia się znacząco.
Stopień tłumienia światła słonecznego i promieniowania kosmicznego zależy od długości fali (częstotliwości) światła. Promieniowanie ultrafioletowe o długości fali mniejszej niż 0,3 mikrona prawie nie przechodzi przez warstwę ozonową (na wysokości około 25 km). Takie promieniowanie jest niebezpieczne dla żywego organizmu, w szczególności dla protoplazmy.
W dzikiej przyrodzie jedynym źródłem energii jest światło, wszystkie rośliny, z wyjątkiem bakterii, poddają się fotosyntezie, tj. syntetyzować substancje organiczne z substancji nieorganicznych (tj. z wody, soli mineralnych i CO 2 - wykorzystując energię promieniowania w procesie asymilacji). Pożywienie wszystkich organizmów jest uzależnione od ziemskich fotosyntezatorów, tj. rośliny chlorofilowe.
Światło jako czynnik środowiskowy dzieli się na ultrafiolet o długości fali 0,40 - 0,75 mikrona i podczerwień o długości fali większej od tych wielkości.
Wpływ tych czynników zależy od właściwości organizmów. Każdy rodzaj organizmu jest przystosowany do takiego lub innego spektrum długości fal światła. Niektóre gatunki organizmów przystosowały się do ultrafioletu, a inne do podczerwieni.
Niektóre organizmy potrafią rozróżnić długość fali. Posiadają specjalne systemy postrzegania światła oraz widzenie barw, które mają ogromne znaczenie w ich życiu. Wiele owadów jest wrażliwych na promieniowanie krótkofalowe, którego ludzie nie dostrzegają. Motyle nocne dobrze odbierają promienie ultrafioletowe. Pszczoły i ptaki dokładnie określają swoje położenie i poruszają się po terenie nawet nocą.
Organizmy również silnie reagują na natężenie światła. Zgodnie z tymi cechami rośliny dzielą się na trzy grupy ekologiczne:
1. Kochające światło, kochające słońce lub heliofity - które są w stanie normalnie rozwijać się tylko pod promieniami słonecznymi.
2. Cienie kochające lub sciophytes to rośliny niższych poziomów lasów i rośliny głębinowe, na przykład konwalie i inne.
Wraz ze spadkiem natężenia światła fotosynteza również zwalnia. Wszystkie żywe organizmy mają progową wrażliwość na natężenie światła, a także na inne czynniki środowiskowe. Na różne organizmy wrażliwość progowa na czynniki środowiskowe nie jest taka sama. Na przykład intensywne światło hamuje rozwój muszek Drosophyll, a nawet powoduje ich śmierć. Nie lubią światła, karaluchów i innych owadów. W większości roślin fotosyntetycznych, przy niskim natężeniu światła, synteza białek jest zahamowana, podczas gdy u zwierząt procesy biosyntezy są zahamowane.
3. Odporne na cień lub fakultatywne heliofity. Rośliny, które dobrze rosną zarówno w cieniu, jak iw świetle. U zwierząt te właściwości organizmów nazywane są kochającymi światło (fotofile), kochającymi cień (fotofobami), euryfobicznymi - stenofobicznymi.
2. Temperatura jako czynnik środowiskowy
Temperatura jest najważniejszym czynnikiem środowiskowym. Temperatura ma ogromny wpływ na wiele aspektów życia organizmów, ich geografię rozmieszczenia, rozmnażanie i inne biologiczne właściwości organizmów, które zależą głównie od temperatury. Zasięg, tj. granice temperatur, w których może istnieć życie, wahają się od około -200°C do +100°C, czasami stwierdza się istnienie bakterii w gorących źródłach o temperaturze 250°C. W rzeczywistości większość organizmów może przetrwać w jeszcze węższym zakresie temperatur.
Niektóre rodzaje mikroorganizmów, głównie bakterie i glony, mogą żyć i namnażać się w gorących źródłach w temperaturach zbliżonych do temperatury wrzenia. Górna granica temperatury dla bakterii z gorących źródeł wynosi około 90°C. Zmienność temperatury jest bardzo ważna z ekologicznego punktu widzenia.
Każdy gatunek jest w stanie żyć tylko w określonym zakresie temperatur, tak zwanych maksymalnych i minimalnych temperaturach śmiertelnych. Poza tymi krytycznymi ekstremalnymi temperaturami, zimnymi lub gorącymi, następuje śmierć organizmu. Gdzieś między nimi jest optymalna temperatura, w której aktywna jest żywotna aktywność wszystkich organizmów, żywej materii jako całości.
Zgodnie z tolerancją organizmów na reżim temperaturowy dzielą się one na eurytermiczne i stenotermiczne, tj. zdolne do wytrzymania szerokich lub wąskich wahań temperatury. Na przykład porosty i wiele bakterii może żyć w różnych temperaturach, a storczyki i inne ciepłolubne rośliny ze stref tropikalnych są ciepłolubne.
Niektóre zwierzęta są w stanie utrzymać stałą temperaturę ciała, niezależnie od temperatury otoczenia. Takie organizmy nazywane są homeotermicznymi. U innych zwierząt temperatura ciała zmienia się w zależności od temperatury otoczenia. Nazywane są poikilothermami. W zależności od sposobu przystosowania się organizmów do reżimu temperaturowego dzieli się je na dwie grupy ekologiczne: kriofile - organizmy przystosowane do zimna, do niskich temperatur; termofile - lub kochający ciepło.
3. Wilgotność jako czynnik środowiskowy
Początkowo wszystkie organizmy były wodne. Po podbiciu ziemi nie utracili zależności od wody. Część integralna wszystkich żywych organizmów to woda. Wilgotność to ilość pary wodnej w powietrzu. Bez wilgoci i wody nie ma życia.
Wilgotność to parametr charakteryzujący zawartość pary wodnej w powietrzu. Wilgotność bezwzględna to ilość pary wodnej w powietrzu i zależy od temperatury i ciśnienia. Ta ilość nazywana jest wilgotnością względną (czyli stosunkiem ilości pary wodnej w powietrzu do ilości pary nasyconej w określonych warunkach temperatury i ciśnienia).
W naturze panuje dobowy rytm wilgotności. Wilgotność zmienia się zarówno w pionie, jak iw poziomie. Czynnik ten, obok światła i temperatury, odgrywa ważną rolę w regulowaniu aktywności organizmów i ich rozmieszczeniu. Wilgotność zmienia również wpływ temperatury.
Suszenie powietrzem jest ważnym czynnikiem środowiskowym. Szczególnie dla organizmów lądowych suszące działanie powietrza ma ogromne znaczenie. Zwierzęta przystosowują się, przenosząc się do obszarów chronionych i są aktywne w nocy.
Rośliny pobierają wodę z gleby i prawie całkowicie (97-99%) odparowują przez liście. Ten proces nazywa się transpiracją. Parowanie chłodzi liście. Dzięki parowaniu jony są transportowane przez glebę do korzeni, transport jonów między komórkami itp.
Dla organizmów lądowych niezbędna jest pewna ilość wilgoci. Wiele z nich do normalnego życia potrzebuje wilgotności względnej 100% i odwrotnie, organizm w stanie normalnym nie może długo żyć w absolutnie suchym powietrzu, ponieważ stale traci wodę. Woda jest istotną częścią żywej materii. Dlatego utrata wody w określonej ilości prowadzi do śmierci.
Rośliny o suchym klimacie przystosowują się do zmian morfologicznych, redukcji organów wegetatywnych, zwłaszcza liści.
Zwierzęta lądowe również się dostosowują. Wielu z nich pije wodę, inni zasysają ją przez powłokę ciała w stanie płynnym lub parowym. Na przykład większość płazów, niektóre owady i roztocza. Większość zwierząt pustynnych nigdy nie pije, zaspokaja swoje potrzeby kosztem wody dostarczanej wraz z pożywieniem. Inne zwierzęta otrzymują wodę w procesie utleniania tłuszczu.
Woda jest niezbędna dla żywych organizmów. Dlatego organizmy rozprzestrzeniają się po całym siedlisku w zależności od swoich potrzeb: organizmy wodne żyją w wodzie stale; hydrofity mogą żyć tylko w bardzo wilgotnym środowisku.
Z punktu widzenia walencji ekologicznej hydrofity i higrofity należą do grupy stenogigerów. Wilgotność ma duży wpływ na funkcje życiowe organizmów, np. wilgotność względna 70% była bardzo korzystna dla dojrzewania polowego i płodności samic. szarańcza wędrowna. Przy korzystnym rozmnażaniu powodują ogromne szkody ekonomiczne w uprawach wielu krajów.
Do oceny ekologicznej rozmieszczenia organizmów stosuje się wskaźnik suchości klimatu. Suchość służy jako czynnik selektywny do ekologicznej klasyfikacji organizmów.
Tak więc, w zależności od charakterystyki wilgotności lokalnego klimatu, gatunki organizmów są rozmieszczone według organizacje ekologiczne:
1. Hydatofity to rośliny wodne.
2. Hydrofity to rośliny lądowo-wodne.
3. Higrofity - rośliny lądowe żyjące w warunkach wysoka wilgotność.
4. Mezofity to rośliny, które rosną ze średnią wilgotnością
5. Kserofity to rośliny rosnące przy niewystarczającej wilgotności. Z kolei dzielą się na: sukulenty - sukulenty (kaktusy); sklerofity to rośliny o wąskich i małych liściach, złożone w kanaliki. Dzielą się również na eukserofity i stipakserofity. Eukserofity to rośliny stepowe. Stipakserofity to grupa wąskolistnych traw darniowych (pióro, kostrzewa, cienkonogie itp.). Z kolei mezofity dzielą się również na mezohygrofity, mezokserofity itp.
Oddając swoją wartość do temperatury, wilgotność jest jednak jednym z głównych czynników środowiskowych. Przez większość historii dzikiej przyrody świat organiczny był reprezentowany wyłącznie przez normy wodne organizmów. Integralną częścią ogromnej większości żywych istot jest woda, a do rozmnażania lub łączenia gamet prawie wszystkie potrzebują środowiska wodnego. Zwierzęta lądowe zmuszone są stworzyć w swoim ciele sztuczne środowisko wodne do zapłodnienia, a to prowadzi do tego, że to ostatnie staje się wewnętrzne.
Wilgotność to ilość pary wodnej w powietrzu. Można go wyrazić w gramach na metr sześcienny.
4. Czynniki edaficzne
Czynniki edaficzne obejmują cały zestaw właściwości fizycznych i chemicznych gleby, które mogą mieć ekologiczny wpływ na organizmy żywe. Odgrywają ważną rolę w życiu organizmów blisko spokrewnionych z glebą. Rośliny są szczególnie uzależnione od czynników edaficznych.
Do głównych właściwości gleby wpływających na życie organizmów należy jej budowa fizyczna, tj. nachylenie, głębokość i granulometria, skład chemiczny samej gleby i krążących w niej substancji - gazy (w tym przypadku konieczne jest poznanie warunków jej napowietrzania), woda, substancje organiczne i mineralne w postaci jonów .
Główną cechą gleby, mającą bardzo ważne zarówno dla roślin, jak i dla zwierząt ryjących się pod ziemią, jest wielkość jego cząstek.
Warunki glebowe gruntu determinowane są czynnikami klimatycznymi. Nawet na małej głębokości w glebie panuje zupełna ciemność, a ta właściwość jest charakterystyczną cechą siedliskową tych gatunków, które unikają światła. W miarę zagłębiania się w glebę wahania temperatury stają się coraz mniej znaczące: dzienne zmiany szybko zanikają, a różnice sezonowe zaczynają się od znanej głębokości. Dzienne różnice temperatur znikają już na głębokości 50 cm, gdy gleba tonie, zawartość tlenu w niej spada, a CO 2 wzrasta. Na znacznej głębokości warunki zbliżają się do warunków beztlenowych, w których żyją niektóre bakterie beztlenowe. Dżdżownice już teraz wolą środowisko o większej zawartości CO 2 niż w atmosferze.
Wilgotność gleby jest bardzo duża ważna cecha szczególnie dla roślin na nim rosnących. Zależy to od wielu czynników: reżimu opadów, głębokości warstwy, a także właściwości fizykochemicznych gleby, której cząstki w zależności od wielkości, zawartości materii organicznej itp. Flora gleb suchych i wilgotnych nie jest taka sama i nie można na tych glebach uprawiać tych samych roślin. Fauna glebowa jest również bardzo wrażliwa na wilgotność gleby i generalnie nie toleruje zbyt dużej suchości. Dobrze znanymi przykładami są dżdżownice i termity. Te ostatnie są czasami zmuszone zaopatrywać swoje kolonie w wodę, tworząc podziemne chodniki na dużych głębokościach. Jednak zbyt duża zawartość wody w glebie zabija duże ilości larw owadów.
Minerały, niezbędne do odżywiania roślin, znajdują się w glebie w postaci jonów rozpuszczonych w wodzie. W glebie można znaleźć co najmniej śladowe ilości ponad 60 pierwiastków chemicznych. CO 2 i azot są obecne w dużych ilościach; zawartość innych, takich jak nikiel czy kobalt, jest bardzo mała. Niektóre jony są trujące dla roślin, inne wręcz przeciwnie, są niezbędne. Stężenie jonów wodorowych w glebie – pH – jest średnio zbliżone do obojętnego. Flora takich gleb jest szczególnie bogata w gatunki. Gleby wapienne i zasolone mają zasadowe pH rzędu 8-9; na torfowiskach torfowych kwaśne pH może spaść do 4.
Niektóre jony mają duże znaczenie ekologiczne. Mogą powodować eliminację wielu gatunków i odwrotnie, przyczyniają się do rozwoju bardzo osobliwych form. Gleby zalegające na wapieniach są bardzo bogate w jon Ca +2; rozwija się na nich specyficzna roślinność, zwana kalcefitem (w górach szarotka; wiele gatunków storczyków). W przeciwieństwie do tej roślinności występuje roślinność kalcefobiczna. Obejmuje kasztany, paprocie paprociowe, większość wrzosów. Roślinność taką nazywa się czasem krzemieniem, ponieważ gleby ubogie w wapń zawierają odpowiednio więcej krzemu. W rzeczywistości ta roślinność nie preferuje bezpośrednio krzemu, ale po prostu unika wapnia. Niektóre zwierzęta mają organiczne zapotrzebowanie na wapń. Wiadomo, że kurczęta przestają składać jaja w twardych skorupkach, jeśli kurnik znajduje się na obszarze o glebie ubogiej w wapń. Strefa wapienna jest licznie zasiedlona przez ślimaki muszlowe (ślimaki), które są tu szeroko reprezentowane pod względem gatunkowym, ale prawie całkowicie znikają na masywach granitowych.
Na glebach bogatych w jony 0,3 rozwija się również specyficzna flora, zwana nitrofilną. Resztki organiczne zawierające azot, które często się na nich znajdują, są rozkładane przez bakterie najpierw na sole amonowe, następnie na azotany, a na końcu na azotany. Rośliny tego typu tworzą np. gęste zarośla w górach w pobliżu pastwisk dla bydła.
Gleba zawiera również materię organiczną powstałą podczas rozkładu martwych roślin i zwierząt. Zawartość tych substancji zmniejsza się wraz ze wzrostem głębokości. Na przykład w lesie ważnym źródłem ich dochodów jest ściółka z opadłych liści, a ściółka gatunków liściastych jest pod tym względem bogatsza niż iglasta. Żywi się organizmami niszczącymi - roślinami saprofitów i zwierzętami saprofitami. Saprofity reprezentowane są głównie przez bakterie i grzyby, ale wśród nich można znaleźć również rośliny wyższe, które utraciły chlorofil w wyniku adaptacji wtórnej. Takie na przykład storczyki.
5. Różne środowiska życia
Według większości autorów badających pochodzenie życia na Ziemi, to środowisko wodne było ewolucyjnym podstawowym środowiskiem życia. Znajdujemy sporo pośrednich potwierdzeń tego stanowiska. Przede wszystkim większość organizmów nie jest zdolna do aktywnego życia bez dostania się wody do organizmu, a przynajmniej bez utrzymywania określonej ilości płynów w organizmie.
Być może główną cechą wyróżniającą środowisko wodne jest jego względny konserwatyzm. Na przykład amplituda wahań temperatury sezonowej lub dobowej w środowisku wodnym jest znacznie mniejsza niż w środowisku ziemia-powietrze. Rzeźba dna, różnica warunków na różnych głębokościach, obecność raf koralowych i tak dalej. stworzyć różnorodne warunki w środowisku wodnym.
Cechy środowiska wodnego wynikają z właściwości fizykochemicznych wody. Tak więc duża gęstość i lepkość wody mają duże znaczenie ekologiczne. Ciężar właściwy wody jest proporcjonalny do ciężaru ciała żywych organizmów. Gęstość wody jest około 1000 razy większa od gęstości powietrza. Dlatego organizmy wodne (zwłaszcza aktywnie poruszające się) napotykają na dużą siłę oporu hydrodynamicznego. Z tego powodu ewolucja wielu grup zwierząt wodnych poszła w kierunku kształtowania się sylwetki i rodzajów ruchu, które zmniejszają opór, co prowadzi do zmniejszenia zużycia energii na pływanie. Tak więc opływowy kształt ciała znajdujemy u przedstawicieli różne grupy organizmy żyjące w wodzie - delfiny (ssaki), ryby kostno-chrzęstne.
Wysoka gęstość wody jest również powodem, dla którego wibracje mechaniczne(drgania) dobrze propagują się w środowisku wodnym. Miało to znaczenie w ewolucji narządów zmysłów, orientacji w przestrzeni i komunikacji między mieszkańcami wody. Czterokrotnie większa niż w powietrzu prędkość dźwięku w środowisku wodnym determinuje wyższą częstotliwość sygnałów echolokacyjnych.
Ze względu na duże zagęszczenie środowiska wodnego jego mieszkańcy pozbawieni są obowiązkowego połączenia z podłożem, co jest charakterystyczne dla form ziemskich i związane jest z siłami grawitacji. Dlatego istnieje cała grupa organizmy wodne (zarówno rośliny, jak i zwierzęta), które istnieją bez obowiązkowego połączenia z dnem lub innym podłożem, „pływając” w słupie wody.
Środowisko gruntowo-powietrzne charakteryzuje się ogromną różnorodnością warunków życia, nisz ekologicznych i zamieszkujących je organizmów.
Głównymi cechami środowiska gruntowo-powietrznego są duża amplituda zmian czynników środowiskowych, niejednorodność środowiska, działanie sił grawitacji oraz niska gęstość powietrza. Kompleks czynników fizjograficznych i klimatycznych tkwiących w określonej strefie naturalnej prowadzi do ewolucyjnego tworzenia morfofizjologicznych adaptacji organizmów do życia w tych warunkach, różnorodnych form życia.
Powietrze atmosferyczne charakteryzuje się niską i zmienną wilgotnością. Okoliczność ta w dużym stopniu ograniczyła (ograniczyła) możliwości opanowania środowiska gruntowo-powietrznego, a także ukierunkowała ewolucję metabolizmu wodno-solnego i budowę narządów oddechowych.
Gleba jest wynikiem działalności żywych organizmów.
Ważną cechą gleby jest również obecność pewnej ilości materii organicznej. Powstaje w wyniku śmierci organizmów i jest częścią ich wydalin (wydzielin).
Warunki siedliskowe glebowe determinują takie właściwości gleby, jak jej napowietrzenie (tj. nasycenie powietrzem), wilgotność (obecność wilgoci), pojemność cieplna i reżim termiczny (dobowe, sezonowe, całoroczne wahania temperatury). Reżim termiczny w porównaniu ze środowiskiem ziemia-powietrze jest bardziej konserwatywny, zwłaszcza na dużych głębokościach. Generalnie gleba charakteryzuje się dość stabilnymi warunkami życia.
Różnice pionowe są również charakterystyczne dla innych właściwości gleby, na przykład przenikanie światła zależy oczywiście od głębokości.
Organizmy glebowe charakteryzują się specyficznymi narządami i typami ruchu (rybiące kończyny u ssaków; zdolność do zmiany grubości ciała; obecność wyspecjalizowanych torebek głowy u niektórych gatunków); kształty ciała (zaokrąglone, w kształcie wilka, w kształcie robaka); trwałe i elastyczne pokrowce; redukcja oczu i zanikanie pigmentów. Wśród mieszkańców gleby szeroko rozwinięta jest saprofagia - jedzenie zwłok innych zwierząt, gnijące szczątki itp.
Wniosek
Wyjście jednego z czynników środowiskowych poza granice wartości minimalnych (progowych) lub maksymalnych (skrajnych) (typowych dla typu strefy tolerancji) grozi śmiercią organizmu nawet przy optymalnej kombinacji innych czynników. Przykładami są: pojawienie się atmosfery tlenowej, epoka lodowcowa, susza, zmiany ciśnienia podczas wynurzania nurków itp.
Każdy czynnik środowiskowy wpływa inaczej różne rodzaje organizmy: optimum dla niektórych może być pessimum dla innych.
Organizmy na powierzchni Ziemi narażone są na przepływ energii, głównie energii słonecznej, a także długofalowego promieniowania cieplnego z ciał kosmicznych. Oba te czynniki determinują warunki klimatyczne środowiska (temperatura, szybkość parowania wody, ruch powietrza i wody).
Temperatura jest najważniejszym czynnikiem środowiskowym. Temperatura ma ogromny wpływ na wiele aspektów życia organizmów, ich geografię rozmieszczenia, rozmnażanie i inne biologiczne właściwości organizmów, które zależą głównie od temperatury.
Suszenie powietrzem jest ważnym czynnikiem środowiskowym. Szczególnie dla organizmów lądowych suszące działanie powietrza ma ogromne znaczenie.
Oddając swoją wartość do temperatury, wilgotność jest jednak jednym z głównych czynników środowiskowych. Przez większość historii dzikiej przyrody świat organiczny był reprezentowany wyłącznie przez normy wodne organizmów.
Czynniki edaficzne obejmują cały zestaw właściwości fizycznych i chemicznych gleby, które mogą mieć ekologiczny wpływ na organizmy żywe. Odgrywają ważną rolę w życiu organizmów blisko spokrewnionych z glebą. Rośliny są szczególnie uzależnione od czynników edaficznych.
Lista wykorzystanej literatury
1. Dedyu I.I. Ekologiczny słownik encyklopedyczny. - Kiszyniów: Wydawnictwo ITU, 1990. - 406 s.
2. Nowikow G.A. Podstawy ekologii ogólnej i ochrony przyrody. - L.: Wydawnictwo Leningrad. un-ta, 1979. - 352 s.
3. Radkiewicz V.A. Ekologia. - Mińsk: Wyższa Szkoła, 1983. - 320 s.
4. Reimers N.F. Ekologia: teoria, prawa, reguły, zasady i hipotezy. -M.: Młoda Rosja, 1994. - 367 s.
5. Riklefs R. Podstawy ekologii ogólnej. - M.: Mir, 1979. - 424 s.
6. Stepanowskich A.S. Ekologia. - Kurgan: GIPP „Zauralje”, 1997. - 616 str.
7. Christoforova N.K. Podstawy ekologii. - Władywostok: Dalnauka, 1999. -517 s.
Wstęp
4. Czynniki edaficzne
5. Różne środowiska życia
Wniosek
Wstęp
Na Ziemi panuje ogromna różnorodność warunków środowiska życia, co zapewnia różnorodność nisz ekologicznych i ich „zasiedlenie”. Jednak pomimo tej różnorodności istnieją cztery jakościowo różne środowiska życia, które mają określony zestaw czynników środowiskowych i dlatego wymagają określonego zestawu. adaptacje. Są to środowiska życia: ziemia-powietrze (ląd); woda; gleba; inne organizmy.
Każdy gatunek jest przystosowany do określonego zestawu warunków środowiskowych - niszy ekologicznej.
Każdy gatunek jest przystosowany do swojego specyficznego środowiska, do określonego pokarmu, drapieżników, temperatury, zasolenia wody i innych elementów świata zewnętrznego, bez których nie może istnieć.
Do istnienia organizmów wymagany jest kompleks czynników. Zapotrzebowanie organizmu na nie jest inne, ale każde w pewnym stopniu ogranicza jego istnienie.
Brak (brak) niektórych czynników środowiskowych może być skompensowany przez inne bliskie (podobne) czynniki. Organizmy nie są „niewolnikami” warunków środowiskowych – do pewnego stopnia same dostosowują się i zmieniają warunki środowiskowe w taki sposób, aby złagodzić brak pewnych czynników.
Braku fizjologicznie niezbędnych czynników (światła, wody, dwutlenku węgla, składników odżywczych) w środowisku nie można zrekompensować (zastąpić) innymi.
1. Światło jako czynnik środowiskowy. Rola światła w życiu organizmów
Światło jest jedną z form energii. Zgodnie z pierwszą zasadą termodynamiki, czyli zasadą zachowania energii, energia może zmieniać się z jednej postaci w drugą. Zgodnie z tym prawem organizmy są systemem termodynamicznym, który nieustannie wymienia energię i materię ze środowiskiem. Organizmy na powierzchni Ziemi narażone są na przepływ energii, głównie energii słonecznej, a także długofalowego promieniowania cieplnego z ciał kosmicznych. Oba te czynniki determinują warunki klimatyczne środowiska (temperatura, szybkość parowania wody, ruch powietrza i wody). Na biosferę pada z kosmosu światło słoneczne o energii 2 cal. na 1 cm2 w 1 min. To tak zwana stała słoneczna. Światło to, przechodząc przez atmosferę, jest osłabione i nie więcej niż 67% jego energii może dotrzeć do powierzchni Ziemi w pogodne południe, tj. 1,34 cal. na cm2 w 1 min. Przechodząc przez zachmurzenie, wodę i roślinność światło słoneczne ulega dalszemu osłabieniu, a rozkład energii w nim w różnych częściach widma zmienia się znacząco.
Stopień tłumienia światła słonecznego i promieniowania kosmicznego zależy od długości fali (częstotliwości) światła. Promieniowanie ultrafioletowe o długości fali mniejszej niż 0,3 mikrona prawie nie przechodzi przez warstwę ozonową (na wysokości około 25 km). Takie promieniowanie jest niebezpieczne dla żywego organizmu, w szczególności dla protoplazmy.
W dzikiej przyrodzie jedynym źródłem energii jest światło, wszystkie rośliny, z wyjątkiem bakterii, poddają się fotosyntezie, tj. syntetyzować substancje organiczne z substancji nieorganicznych (tj. z wody, soli mineralnych i CO 2 - wykorzystując energię promieniowania w procesie asymilacji). Pożywienie wszystkich organizmów jest uzależnione od ziemskich fotosyntezatorów, tj. rośliny chlorofilowe.
Światło jako czynnik środowiskowy dzieli się na ultrafiolet o długości fali 0,40 - 0,75 mikrona i podczerwień o długości fali większej od tych wielkości.
Wpływ tych czynników zależy od właściwości organizmów. Każdy rodzaj organizmu jest przystosowany do takiego lub innego spektrum długości fal światła. Niektóre gatunki organizmów przystosowały się do ultrafioletu, a inne do podczerwieni.
Niektóre organizmy potrafią rozróżnić długość fali. Posiadają specjalne systemy postrzegania światła oraz widzenie barw, które mają ogromne znaczenie w ich życiu. Wiele owadów jest wrażliwych na promieniowanie krótkofalowe, którego ludzie nie dostrzegają. Motyle nocne dobrze odbierają promienie ultrafioletowe. Pszczoły i ptaki dokładnie określają swoje położenie i poruszają się po terenie nawet nocą.
Organizmy również silnie reagują na natężenie światła. Zgodnie z tymi cechami rośliny dzielą się na trzy grupy ekologiczne:
1. Kochające światło, kochające słońce lub heliofity - które są w stanie normalnie rozwijać się tylko pod promieniami słonecznymi.
2. Cienie kochające lub sciophytes to rośliny niższych poziomów lasów i rośliny głębinowe, na przykład konwalie i inne.
Wraz ze spadkiem natężenia światła fotosynteza również zwalnia. Wszystkie żywe organizmy mają progową wrażliwość na natężenie światła, a także na inne czynniki środowiskowe. Różne organizmy mają różną wrażliwość progową na czynniki środowiskowe. Na przykład intensywne światło hamuje rozwój muszek Drosophyll, a nawet powoduje ich śmierć. Nie lubią światła, karaluchów i innych owadów. W większości roślin fotosyntetycznych, przy niskim natężeniu światła, synteza białek jest zahamowana, podczas gdy u zwierząt procesy biosyntezy są zahamowane.
3. Odporne na cień lub fakultatywne heliofity. Rośliny, które dobrze rosną zarówno w cieniu, jak iw świetle. U zwierząt te właściwości organizmów nazywane są kochającymi światło (fotofile), kochającymi cień (fotofobami), euryfobicznymi - stenofobicznymi.
2. Temperatura jako czynnik środowiskowy
Temperatura jest najważniejszym czynnikiem środowiskowym. Temperatura ma ogromny wpływ na wiele aspektów życia organizmów, ich geografię rozmieszczenia, rozmnażanie i inne biologiczne właściwości organizmów, które zależą głównie od temperatury. Zasięg, tj. granice temperatur, w których może istnieć życie, wahają się od około -200°C do +100°C, czasami stwierdza się istnienie bakterii w gorących źródłach o temperaturze 250°C. W rzeczywistości większość organizmów może przetrwać w jeszcze węższym zakresie temperatur.
Niektóre rodzaje mikroorganizmów, głównie bakterie i glony, mogą żyć i namnażać się w gorących źródłach w temperaturach zbliżonych do temperatury wrzenia. Górna granica temperatury dla bakterii z gorących źródeł wynosi około 90°C. Zmienność temperatury jest bardzo ważna z ekologicznego punktu widzenia.
Każdy gatunek jest w stanie żyć tylko w określonym zakresie temperatur, tak zwanych maksymalnych i minimalnych temperaturach śmiertelnych. Poza tymi krytycznymi ekstremalnymi temperaturami, zimnymi lub gorącymi, następuje śmierć organizmu. Gdzieś między nimi jest optymalna temperatura, w której aktywna jest żywotna aktywność wszystkich organizmów, żywej materii jako całości.
Zgodnie z tolerancją organizmów na reżim temperaturowy dzieli się je na eurytermiczne i ciepłolubne, tj. zdolne do wytrzymania szerokich lub wąskich wahań temperatury. Na przykład porosty i wiele bakterii może żyć w różnych temperaturach, storczyki i inne rośliny ciepłolubne strefy tropikalne - są ciepłolubne.
Niektóre zwierzęta są w stanie utrzymać stałą temperaturę ciała, niezależnie od temperatury otoczenia. Takie organizmy nazywane są homeotermicznymi. U innych zwierząt temperatura ciała zmienia się w zależności od temperatury otoczenia. Nazywane są poikilothermami. W zależności od sposobu przystosowania się organizmów do reżimu temperaturowego dzieli się je na dwie grupy ekologiczne: kriofile - organizmy przystosowane do zimna, do niskich temperatur; termofile - lub kochający ciepło.
3. Wilgotność jako czynnik środowiskowy
Początkowo wszystkie organizmy były wodne. Po podbiciu ziemi nie utracili zależności od wody. Woda jest integralną częścią wszystkich żywych organizmów. Wilgotność to ilość pary wodnej w powietrzu. Bez wilgoci i wody nie ma życia.
Wilgotność to parametr charakteryzujący zawartość pary wodnej w powietrzu. Wilgotność bezwzględna to ilość pary wodnej w powietrzu i zależy od temperatury i ciśnienia. Ta ilość nazywana jest wilgotnością względną (czyli stosunkiem ilości pary wodnej w powietrzu do ilości pary nasyconej w określonych warunkach temperatury i ciśnienia).
W naturze panuje dobowy rytm wilgotności. Wilgotność zmienia się zarówno w pionie, jak iw poziomie. Czynnik ten, obok światła i temperatury, odgrywa ważną rolę w regulowaniu aktywności organizmów i ich rozmieszczeniu. Wilgotność zmienia również wpływ temperatury.
Suszenie powietrzem jest ważnym czynnikiem środowiskowym. Szczególnie dla organizmów lądowych suszące działanie powietrza ma ogromne znaczenie. Zwierzęta przystosowują się, przenosząc się do obszarów chronionych i są aktywne w nocy.
Rośliny pobierają wodę z gleby i prawie całkowicie (97-99%) odparowują przez liście. Ten proces nazywa się transpiracją. Parowanie chłodzi liście. Dzięki parowaniu jony są transportowane przez glebę do korzeni, transport jonów między komórkami itp.
Dla organizmów lądowych niezbędna jest pewna ilość wilgoci. Wiele z nich do normalnego życia potrzebuje wilgotności względnej 100% i odwrotnie, organizm w stanie normalnym nie może długo żyć w absolutnie suchym powietrzu, ponieważ stale traci wodę. Woda jest istotną częścią żywej materii. Dlatego utrata wody w określonej ilości prowadzi do śmierci.
Rośliny o suchym klimacie przystosowują się do zmian morfologicznych, redukcji organów wegetatywnych, zwłaszcza liści.
Zwierzęta lądowe również się dostosowują. Wielu z nich pije wodę, inni zasysają ją przez powłokę ciała w stanie płynnym lub parowym. Na przykład większość płazów, niektóre owady i roztocza. Większość zwierząt pustynnych nigdy nie pije, zaspokaja swoje potrzeby kosztem wody dostarczanej wraz z pożywieniem. Inne zwierzęta otrzymują wodę w procesie utleniania tłuszczu.
Woda jest niezbędna dla żywych organizmów. Dlatego organizmy rozprzestrzeniają się po całym siedlisku w zależności od swoich potrzeb: organizmy wodne żyją w wodzie stale; hydrofity mogą żyć tylko w bardzo wilgotnym środowisku.
Z punktu widzenia walencji ekologicznej hydrofity i higrofity należą do grupy stenogigerów. Wilgotność ma duży wpływ na funkcje życiowe organizmów, na przykład wilgotność względna 70% była bardzo korzystna dla dojrzewania polowego i płodności samic wędrownych szarańczy. Przy korzystnym rozmnażaniu powodują ogromne szkody ekonomiczne w uprawach wielu krajów.
Do oceny ekologicznej rozmieszczenia organizmów stosuje się wskaźnik suchości klimatu. Suchość służy jako czynnik selektywny do ekologicznej klasyfikacji organizmów.
Tak więc, w zależności od charakterystyki wilgotności lokalnego klimatu, gatunki organizmów są podzielone na grupy ekologiczne:
1. Hydatofity to rośliny wodne.
2. Hydrofity to rośliny lądowo-wodne.
3. Higrofity - rośliny lądowe żyjące w warunkach dużej wilgotności.
4. Mezofity to rośliny, które rosną ze średnią wilgotnością
5. Kserofity to rośliny rosnące przy niewystarczającej wilgotności. Z kolei dzielą się na: sukulenty - sukulenty (kaktusy); sklerofity to rośliny o wąskich i małych liściach, złożone w kanaliki. Dzielą się również na eukserofity i stipakserofity. Eukserofity to rośliny stepowe. Stipakserofity to grupa wąskolistnych traw darniowych (pióro, kostrzewa, cienkonogie itp.). Z kolei mezofity dzielą się również na mezohygrofity, mezokserofity itp.
Oddając swoją wartość do temperatury, wilgotność jest jednak jednym z głównych czynników środowiskowych. Przez większość historii dzikiej przyrody świat organiczny był reprezentowany wyłącznie przez normy wodne organizmów. Integralną częścią ogromnej większości żywych istot jest woda, a do rozmnażania lub łączenia gamet prawie wszystkie potrzebują środowiska wodnego. Zwierzęta lądowe zmuszone są stworzyć w swoim ciele sztuczne środowisko wodne do zapłodnienia, a to prowadzi do tego, że to ostatnie staje się wewnętrzne.
Wilgotność to ilość pary wodnej w powietrzu. Można go wyrazić w gramach na metr sześcienny.
4. Czynniki edaficzne
Do głównych właściwości gleby wpływających na życie organizmów należy jej budowa fizyczna, tj. nachylenie, głębokość i granulometria, skład chemiczny samej gleby i krążących w niej substancji - gazy (w tym przypadku konieczne jest poznanie warunków jej napowietrzania), woda, substancje organiczne i mineralne w postaci jonów .
Główną cechą gleby, która ma duże znaczenie zarówno dla roślin, jak i zwierząt ryjących, jest wielkość jej cząstek.
Warunki glebowe gruntu determinowane są czynnikami klimatycznymi. Nawet na małej głębokości w glebie panuje zupełna ciemność, a ta właściwość jest charakterystyczną cechą siedliskową tych gatunków, które unikają światła. W miarę zagłębiania się w glebę wahania temperatury stają się coraz mniej znaczące: dzienne zmiany szybko zanikają, a różnice sezonowe zaczynają się od znanej głębokości. Dzienne różnice temperatur znikają już na głębokości 50 cm, gdy gleba tonie, zawartość tlenu w niej spada, a CO 2 wzrasta. Na znacznej głębokości warunki zbliżają się do warunków beztlenowych, w których żyją niektóre bakterie beztlenowe. Dżdżownice już teraz wolą środowisko o większej zawartości CO 2 niż w atmosferze.
Wilgotność gleby jest niezwykle ważną cechą, zwłaszcza dla rosnących na niej roślin. Zależy to od wielu czynników: reżimu opadów, głębokości warstwy, a także właściwości fizykochemicznych gleby, której cząstki w zależności od wielkości, zawartości materii organicznej itp. Flora gleb suchych i wilgotnych nie jest taka sama i nie można na tych glebach uprawiać tych samych roślin. Fauna glebowa jest również bardzo wrażliwa na wilgotność gleby i generalnie nie toleruje zbyt dużej suchości. Dobrze znanymi przykładami są dżdżownice i termity. Te ostatnie są czasami zmuszone zaopatrywać swoje kolonie w wodę, tworząc podziemne chodniki na dużych głębokościach. Jednak zbyt duża zawartość wody w glebie zabija duże ilości larw owadów.
Minerały niezbędne do odżywiania roślin znajdują się w glebie w postaci jonów rozpuszczonych w wodzie. W glebie można znaleźć co najmniej śladowe ilości ponad 60 pierwiastków chemicznych. CO 2 i azot są obecne w dużych ilościach; zawartość innych, takich jak nikiel czy kobalt, jest bardzo mała. Niektóre jony są trujące dla roślin, inne wręcz przeciwnie, są niezbędne. Stężenie jonów wodorowych w glebie – pH – jest średnio zbliżone do obojętnego. Flora takich gleb jest szczególnie bogata w gatunki. Gleby wapienne i zasolone mają zasadowe pH rzędu 8-9; na torfowiskach torfowych kwaśne pH może spaść do 4.
Niektóre jony mają duże znaczenie ekologiczne. Mogą powodować eliminację wielu gatunków i odwrotnie, przyczyniają się do rozwoju bardzo osobliwych form. Gleby zalegające na wapieniach są bardzo bogate w jon Ca +2; rozwija się na nich specyficzna roślinność, zwana kalcefitem (w górach szarotka; wiele gatunków storczyków). W przeciwieństwie do tej roślinności występuje roślinność kalcefobiczna. Obejmuje kasztany, paprocie paprociowe, większość wrzosów. Roślinność taką nazywa się czasem krzemieniem, ponieważ gleby ubogie w wapń zawierają odpowiednio więcej krzemu. W rzeczywistości ta roślinność nie preferuje bezpośrednio krzemu, ale po prostu unika wapnia. Niektóre zwierzęta mają organiczne zapotrzebowanie na wapń. Wiadomo, że kurczęta przestają składać jaja w twardych skorupkach, jeśli kurnik znajduje się na obszarze o glebie ubogiej w wapń. Strefa wapienna jest licznie zasiedlona przez ślimaki muszlowe (ślimaki), które są tu szeroko reprezentowane pod względem gatunkowym, ale prawie całkowicie znikają na masywach granitowych.
Na glebach bogatych w jony 0,3 rozwija się również specyficzna flora, zwana nitrofilną. Resztki organiczne zawierające azot, które często się na nich znajdują, są rozkładane przez bakterie najpierw na sole amonowe, następnie na azotany, a na końcu na azotany. Rośliny tego typu tworzą np. gęste zarośla w górach w pobliżu pastwisk dla bydła.
Gleba zawiera również materię organiczną powstałą podczas rozkładu martwych roślin i zwierząt. Zawartość tych substancji zmniejsza się wraz ze wzrostem głębokości. Na przykład w lesie ważnym źródłem ich dochodów jest ściółka z opadłych liści, a ściółka gatunków liściastych jest pod tym względem bogatsza niż iglasta. Żywi się organizmami niszczącymi - roślinami saprofitów i zwierzętami saprofitami. Saprofity reprezentowane są głównie przez bakterie i grzyby, ale wśród nich można znaleźć również rośliny wyższe, które utraciły chlorofil w wyniku adaptacji wtórnej. Takie na przykład storczyki.
5. Różne środowiska życia
Według większości autorów badających pochodzenie życia na Ziemi, to środowisko wodne było ewolucyjnym podstawowym środowiskiem życia. Znajdujemy sporo pośrednich potwierdzeń tego stanowiska. Przede wszystkim większość organizmów nie jest zdolna do aktywnego życia bez dostania się wody do organizmu, a przynajmniej bez utrzymywania określonej ilości płynów w organizmie.
Być może główną cechą wyróżniającą środowisko wodne jest jego względny konserwatyzm. Na przykład amplituda wahań temperatury sezonowej lub dobowej w środowisku wodnym jest znacznie mniejsza niż w środowisku ziemia-powietrze. Rzeźba dna, różnica warunków na różnych głębokościach, obecność raf koralowych i tak dalej. stworzyć różnorodne warunki w środowisku wodnym.
Cechy środowiska wodnego wynikają z: fizyczne i chemiczne właściwości woda. Tak więc duża gęstość i lepkość wody mają duże znaczenie ekologiczne. Ciężar właściwy wody jest proporcjonalny do ciężaru ciała żywych organizmów. Gęstość wody jest około 1000 razy większa od gęstości powietrza. Dlatego organizmy wodne (zwłaszcza aktywnie poruszające się) napotykają na dużą siłę oporu hydrodynamicznego. Z tego powodu ewolucja wielu grup zwierząt wodnych poszła w kierunku kształtowania się sylwetki i rodzajów ruchu, które zmniejszają opór, co prowadzi do zmniejszenia zużycia energii na pływanie. Tak więc opływowy kształt ciała występuje u przedstawicieli różnych grup organizmów żyjących w wodzie - delfinów (ssaków), ryb kostnych i chrzęstnych.
Wysoka gęstość wody powoduje również, że drgania mechaniczne (wibracje) dobrze propagują się w środowisku wodnym. Miało to znaczenie w ewolucji narządów zmysłów, orientacji w przestrzeni i komunikacji między mieszkańcami wody. Czterokrotnie większa niż w powietrzu prędkość dźwięku w środowisku wodnym determinuje wyższą częstotliwość sygnałów echolokacyjnych.
Ze względu na duże zagęszczenie środowiska wodnego jego mieszkańcy pozbawieni są obowiązkowego połączenia z podłożem, co jest charakterystyczne dla form ziemskich i związane jest z siłami grawitacji. Istnieje więc cała grupa organizmów wodnych (zarówno roślin, jak i zwierząt), które istnieją bez obowiązkowego połączenia z dnem lub innym podłożem, „pływając” w słupie wody.
Przewodnictwo elektryczne otworzyło możliwość ewolucyjnego tworzenia elektrycznych narządów zmysłów, obrony i ataku.
Środowisko gruntowo-powietrzne charakteryzuje się ogromną różnorodnością warunków życia, nisz ekologicznych i zamieszkujących je organizmów.
Głównymi cechami środowiska gruntowo-powietrznego są duża amplituda zmian czynników środowiskowych, niejednorodność środowiska, działanie sił grawitacji oraz niska gęstość powietrza. Kompleks czynników fizjograficznych i klimatycznych tkwiących w określonej strefie naturalnej prowadzi do ewolucyjnego tworzenia morfofizjologicznych adaptacji organizmów do życia w tych warunkach, różnorodnych form życia.
Powietrze atmosferyczne charakteryzuje się niską i zmienną wilgotnością. Okoliczność ta w dużym stopniu ograniczyła (ograniczyła) możliwości opanowania środowiska gruntowo-powietrznego, a także ukierunkowała ewolucję metabolizmu wodno-solnego i budowę narządów oddechowych.
Gleba jest wynikiem działalności żywych organizmów.
Ważną cechą gleby jest również obecność pewnej ilości materii organicznej. Powstaje w wyniku śmierci organizmów i jest częścią ich wydalin (wydzielin).
Warunki siedliskowe glebowe determinują takie właściwości gleby, jak jej napowietrzenie (tj. nasycenie powietrzem), wilgotność (obecność wilgoci), pojemność cieplna i reżim termiczny (dobowe, sezonowe, całoroczne wahania temperatury). Reżim termiczny w porównaniu ze środowiskiem ziemia-powietrze jest bardziej konserwatywny, zwłaszcza na dużych głębokościach. Generalnie gleba charakteryzuje się dość stabilnymi warunkami życia.
Różnice pionowe są również charakterystyczne dla innych właściwości gleby, na przykład przenikanie światła zależy oczywiście od głębokości.
Organizmy glebowe charakteryzują się specyficznymi narządami i typami ruchu (rybiące kończyny u ssaków; zdolność do zmiany grubości ciała; obecność wyspecjalizowanych torebek głowy u niektórych gatunków); kształty ciała (zaokrąglone, w kształcie wilka, w kształcie robaka); trwałe i elastyczne pokrowce; redukcja oczu i zanikanie pigmentów. Wśród mieszkańców gleby szeroko rozwinięta jest saprofagia - jedzenie zwłok innych zwierząt, gnijące szczątki itp.
Wniosek
Wyjście jednego z czynników środowiskowych poza granice wartości minimalnych (progowych) lub maksymalnych (skrajnych) (typowych dla typu strefy tolerancji) grozi śmiercią organizmu nawet przy optymalnej kombinacji innych czynników. Przykładami są: pojawienie się atmosfery tlenowej, epoka lodowcowa, susza, zmiany ciśnienia podczas wynurzania nurków itp.
Każdy czynnik środowiskowy w różny sposób wpływa na różne typy organizmów: optimum dla niektórych może być pessimum dla innych.
Organizmy na powierzchni Ziemi narażone są na przepływ energii, głównie energii słonecznej, a także długofalowego promieniowania cieplnego z ciał kosmicznych. Oba te czynniki determinują warunki klimatyczne środowiska (temperatura, szybkość parowania wody, ruch powietrza i wody).
Temperatura jest najważniejszym czynnikiem środowiskowym. Temperatura ma ogromny wpływ na wiele aspektów życia organizmów, ich geografię rozmieszczenia, rozmnażanie i inne biologiczne właściwości organizmów, które zależą głównie od temperatury.
Suszenie powietrzem jest ważnym czynnikiem środowiskowym. Szczególnie dla organizmów lądowych suszące działanie powietrza ma ogromne znaczenie.
Oddając swoją wartość do temperatury, wilgotność jest jednak jednym z głównych czynników środowiskowych. Przez większość historii dzikiej przyrody świat organiczny był reprezentowany wyłącznie przez normy wodne organizmów.
Czynniki edaficzne obejmują cały zestaw właściwości fizycznych i chemicznych gleby, które mogą mieć ekologiczny wpływ na organizmy żywe. Odgrywają ważną rolę w życiu organizmów blisko spokrewnionych z glebą. Rośliny są szczególnie uzależnione od czynników edaficznych.
Lista wykorzystanej literatury
1. Dedyu I.I. Ekologiczny słownik encyklopedyczny. - Kiszyniów: Wydawnictwo ITU, 1990. - 406 s.
2. Nowikow G.A. Podstawy ekologii ogólnej i ochrony przyrody. - L.: Wydawnictwo Leningrad. un-ta, 1979. - 352 s.
3. Radkiewicz V.A. Ekologia. - Mińsk: Wyższa Szkoła, 1983. - 320 s.
4. Reimers N.F. Ekologia: teoria, prawa, reguły, zasady i hipotezy. -M.: Młoda Rosja, 1994. - 367 s.
5. Riklefs R. Podstawy ekologii ogólnej. - M.: Mir, 1979. - 424 s.
6. Stepanowskich A.S. Ekologia. - Kurgan: GIPP „Zauralje”, 1997. - 616 str.
7. Christoforova N.K. Podstawy ekologii. - Władywostok: Dalnauka, 1999. -517 s.
Więcej z działu Ekologia:
- Zajęcia: Wpływ czynników chemicznych, fizycznych i biologicznych w trakcie procesu technologicznego na środowisko i zarządzanie zdrowiem człowieka
- Test: Strategia pokonywania globalnych zagrożeń środowiskowych
- Streszczenie: Wykorzystanie technologii informatycznych do diagnozowania i poprawy stanu środowiska i zdrowia człowieka
Początkowo wszystkie organizmy były wodne. Po podbiciu ziemi nie utracili zależności od wody. Woda jest integralną częścią wszystkich żywych organizmów. Wilgotność to ilość pary wodnej w powietrzu. Bez wilgoci i wody nie ma życia.
Wilgotność to parametr charakteryzujący zawartość pary wodnej w powietrzu. Wilgotność bezwzględna to ilość pary wodnej w powietrzu i zależy od temperatury i ciśnienia. Ta ilość nazywana jest wilgotnością względną (czyli stosunkiem ilości pary wodnej w powietrzu do ilości pary nasyconej w określonych warunkach temperatury i ciśnienia).
W naturze panuje dobowy rytm wilgotności. Wilgotność zmienia się zarówno w pionie, jak iw poziomie. Czynnik ten, obok światła i temperatury, odgrywa ważną rolę w regulowaniu aktywności organizmów i ich rozmieszczeniu. Wilgotność zmienia również wpływ temperatury.
Suszenie powietrzem jest ważnym czynnikiem środowiskowym. Szczególnie dla organizmów lądowych suszące działanie powietrza ma ogromne znaczenie. Zwierzęta przystosowują się, przenosząc się do obszarów chronionych i są aktywne w nocy.
Rośliny pobierają wodę z gleby i prawie całkowicie (97-99%) odparowują przez liście. Ten proces nazywa się transpiracją. Parowanie chłodzi liście. Dzięki parowaniu jony są transportowane przez glebę do korzeni, transport jonów między komórkami itp.
Dla organizmów lądowych niezbędna jest pewna ilość wilgoci. Wiele z nich do normalnego życia potrzebuje wilgotności względnej 100% i odwrotnie, organizm w stanie normalnym nie może długo żyć w absolutnie suchym powietrzu, ponieważ stale traci wodę. Woda jest istotną częścią żywej materii. Dlatego utrata wody w określonej ilości prowadzi do śmierci.
Rośliny o suchym klimacie przystosowują się do zmian morfologicznych, redukcji organów wegetatywnych, zwłaszcza liści.
Zwierzęta lądowe również się dostosowują. Wielu z nich pije wodę, inni zasysają ją przez powłokę ciała w stanie płynnym lub parowym. Na przykład większość płazów, niektóre owady i roztocza. Większość zwierząt pustynnych nigdy nie pije, zaspokaja swoje potrzeby kosztem wody dostarczanej wraz z pożywieniem. Inne zwierzęta otrzymują wodę w procesie utleniania tłuszczu.
Woda jest niezbędna dla żywych organizmów. Dlatego organizmy rozprzestrzeniają się po całym siedlisku w zależności od swoich potrzeb: organizmy wodne żyją w wodzie stale; hydrofity mogą żyć tylko w bardzo wilgotnym środowisku.
Z punktu widzenia walencji ekologicznej hydrofity i higrofity należą do grupy stenogigerów. Wilgotność ma duży wpływ na funkcje życiowe organizmów, na przykład wilgotność względna 70% była bardzo korzystna dla dojrzewania polowego i płodności samic wędrownych szarańczy. Przy korzystnym rozmnażaniu powodują ogromne szkody ekonomiczne w uprawach wielu krajów.
Do oceny ekologicznej rozmieszczenia organizmów stosuje się wskaźnik suchości klimatu. Suchość służy jako czynnik selektywny do ekologicznej klasyfikacji organizmów.
Tak więc, w zależności od charakterystyki wilgotności lokalnego klimatu, gatunki organizmów są podzielone na grupy ekologiczne:
- 1. Hydatofity to rośliny wodne.
- 2. Hydrofity to rośliny lądowo-wodne.
- 3. Higrofity - rośliny lądowe żyjące w warunkach dużej wilgotności.
- 4. Mezofity to rośliny, które rosną ze średnią wilgotnością
- 5. Kserofity to rośliny rosnące przy niewystarczającej wilgotności. Z kolei dzielą się na: sukulenty - sukulenty (kaktusy); sklerofity to rośliny o wąskich i małych liściach, złożone w kanaliki. Dzielą się również na eukserofity i stipakserofity. Eukserofity to rośliny stepowe. Stipakserofity to grupa wąskolistnych traw darniowych (pióro, kostrzewa, cienkonogie itp.). Z kolei mezofity dzielą się również na mezohygrofity, mezokserofity itp.
Oddając swoją wartość do temperatury, wilgotność jest jednak jednym z głównych czynników środowiskowych. Przez większość historii dzikiej przyrody świat organiczny był reprezentowany wyłącznie przez normy wodne organizmów. Integralną częścią ogromnej większości żywych istot jest woda, a do rozmnażania lub łączenia gamet prawie wszystkie potrzebują środowiska wodnego. Zwierzęta lądowe zmuszone są stworzyć w swoim ciele sztuczne środowisko wodne do zapłodnienia, a to prowadzi do tego, że to ostatnie staje się wewnętrzne.
Wilgotność to ilość pary wodnej w powietrzu. Można go wyrazić w gramach na metr sześcienny.