przystosowane rośliny. Główne etapy ewolucji roślin i zwierząt. Rodzaje roślin a reżim wodny
Przeczytaj także
Teraz, gdy poznaliśmy wyróżniające cechy czterech głównych grup roślin, a mianowicie mszaków, paproci, nagonasiennych i okrytonasiennych (rośliny kwitnące), łatwiej jest nam wyobrazić sobie postęp ewolucyjny, jaki dokonały się u roślin w procesie przystosowanie się do życia na lądzie.
Problemy
Być może najtrudniejszym problemem, który trzeba było jakoś przezwyciężyć, aby przejść z wodnego stylu życia na naziemny, był problem odwodnienie. Każda roślina, która nie jest chroniona w ten czy inny sposób, na przykład nie pokryta woskowym naskórkiem, bardzo szybko wyschnie i niewątpliwie umrze. Nawet jeśli ta trudność zostanie przezwyciężona, inne nierozwiązane problemy pozostają. A przede wszystkim pytanie, jak skutecznie przeprowadzić rozmnażanie płciowe. W pierwszych roślinach rozmnażały się gamety męskie, zdolne do zbliżenia się do gamet żeńskich tylko przez pływanie w wodzie.
Zazwyczaj uważa się, że pierwsze rośliny, które opanowały ziemię, pochodziły od zielonych alg, u niektórych najbardziej zaawansowanych ewolucyjnie przedstawicieli, których pojawiły się narządy rozrodcze, a mianowicie archegonia (samica) i antheridia (samiec); w tych narządach gamety były ukryte, a co za tym idzie chronione. Ta okoliczność i szereg innych dobrze zdefiniowanych urządzeń, które pomagają uniknąć wysychania, pozwoliły niektórym przedstawicielom zielonych glonów przejąć ziemię.
Jednym z najważniejszych trendów ewolucyjnych w roślinach jest ich stopniowo rosnąca niezależność od wody.
Poniżej wymieniono główne trudności związane z przejściem od bytu wodnego do ziemskiego.
- Odwodnienie. Powietrze jest środkiem osuszającym, a woda jest niezbędna do życia z różnych powodów (sekcja 3.1.2). Dlatego istnieje zapotrzebowanie na urządzenia do pozyskiwania i magazynowania wody.
- Reprodukcja. Delikatne komórki płciowe muszą być chronione, a ruchliwe gamety męskie (plemniki) mogą spotkać gamety żeńskie tylko w wodzie.
- Wspierać się. W przeciwieństwie do wody powietrze nie wspiera roślin.
- Żywność. Rośliny potrzebują światła i dwutlenku węgla (CO 2 ) do fotosyntezy, więc przynajmniej część rośliny musi znajdować się nad ziemią. Jednak sole mineralne i woda znajdują się w glebie lub na jej powierzchni i aby skutecznie wykorzystać te substancje, część rośliny musi znajdować się w ziemi i rosnąć w ciemności.
- Wymiana gazowa. W przypadku fotosyntezy i oddychania konieczne jest, aby wymiana dwutlenku węgla i tlenu zachodziła nie z otaczającym roztworem, ale z atmosferą.
- czynniki środowiskowe. Woda, zwłaszcza gdy jest jej tak dużo, jak na przykład w jeziorze lub oceanie, zapewnia wysoką stałość warunków środowiskowych. Siedlisko lądowe natomiast zdecydowanie bardziej charakteryzuje się zmiennością tak ważnych czynników jak temperatura, natężenie światła, stężenie jonów i pH.
Wątrobowce i mchy
Mchy są dobrze przystosowane do rozprzestrzeniania się zarodników w warunkach lądowych: zależy to od wysuszenia skrzynki i rozproszenia małych, lekkich zarodników przez wiatr. Jednak te rośliny są nadal zależne od wody z następujących powodów.
- Do rozmnażania potrzebują wody, ponieważ plemniki muszą dopłynąć do archegonium. Rośliny te wykształciły adaptacje, które pozwalają im uwalniać plemniki tylko w wilgotnym środowisku, ponieważ tylko w takim środowisku pylniki otwierają się. Rośliny te częściowo przystosowały się do życia naziemnego, gdyż ich gamety tworzą się w strukturach ochronnych - antheridia i archegonia.
- Nie mają specjalnych tkanek podporowych, dlatego wzrost rośliny w górę jest ograniczony.
- Mszaki nie mają korzeni, które mogą wnikać głęboko w podłoże i mogą żyć tylko tam, gdzie na powierzchni gleby lub w jej górnych warstwach jest wystarczająca ilość wilgoci i soli mineralnych. Mają jednak ryzoidy, którymi przyczepiają się do ziemi; jest to jedna z adaptacji do życia na stałym podłożu.
2.4. Wątrobowce i mchy są często nazywane płazami (płazami) świata roślin. Wyjaśnij krótko dlaczego.
paprocie
2.5. Paprocie lepiej przystosowały się do życia na lądzie niż wątrobowce i mchy. Jak to jest pokazane?
2.6. Jakie są ważne cechy mchów, paproci i wątrobowców, które są słabo przystosowane do życia na lądzie?
Rośliny nasienne - iglaste i kwitnące
Jedna z głównych trudności, z jakimi borykają się rośliny na lądzie, jest związana z wrażliwością generacji gametofitów. Na przykład u paproci gametofit jest delikatną naroślą, która wytwarza męskie gamety (plemniki), które potrzebują wody, aby dotrzeć do jaja. Jednak w roślinach nasiennych gametofit jest chroniony i znacznie zmniejszony.
Rośliny nasienne mają trzy ważne zalety: po pierwsze, są niejednorodne; po drugie, pojawienie się niepływających gamet męskich i, po trzecie, tworzenie nasion.
RÓŻNORODNOŚĆ I NIEPŁYWAJĄCE GRY MĘSKIE.
Ryż. 2.34. Uogólniony schemat cyklu życia roślin, odzwierciedlający przemianę pokoleń. Zwróć uwagę na obecność stadiów haploidalnych (n) i diploidalnych (2n). Gametofit jest zawsze haploidalny i zawsze tworzy gamety przez podział mitotyczny. Sporofit jest zawsze diploidalny i zawsze tworzy zarodniki w wyniku podziału mejotycznego.
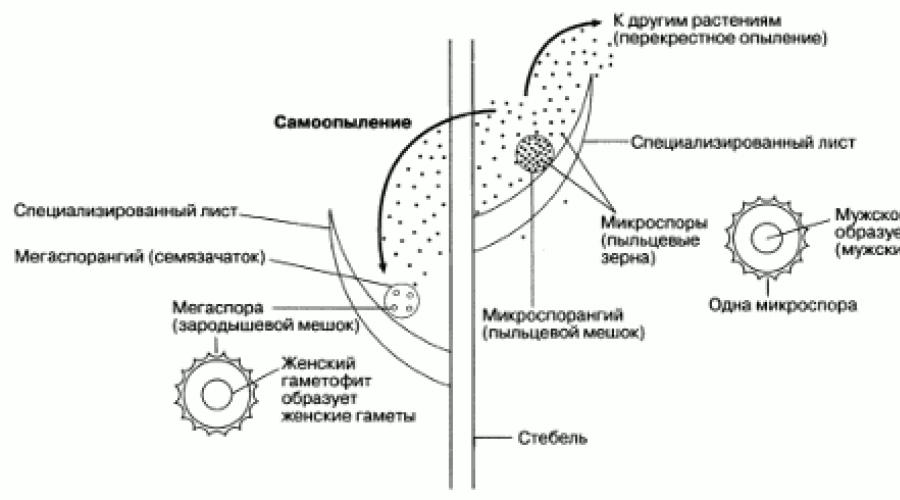
Bardzo ważną rolę w ewolucji roślin odegrało pojawienie się niektórych paproci i ich bliskich krewnych, które tworzą zarodniki dwóch typów. Zjawisko to nazywa się niejednorodność, a rośliny są heterospory. Wszystko rośliny nasienne są heterospory. Tworzą duże zarodniki zwane megaspory, w zarodniach jednego typu (megasporangia) i małych zarodnikach, zwanych mikrosporangiami, w innym typie zarodni (mikrosporangia). Kiełkujące zarodniki tworzą gametofity (ryc. 2.34). Megaspory rozwijają się w żeńskie gametofity, mikrospory w męskie. W roślinach nasiennych gametofity utworzone przez megaspory i mikrospory są bardzo małe i nigdy nie są uwalniane z zarodników. W ten sposób gametofity są chronione przed wysychaniem, co jest ważnym osiągnięciem ewolucyjnym. Jednak plemniki z gametofitu męskiego nadal muszą przenieść się do gametofitu żeńskiego, co znacznie ułatwia dyspersja mikrospor. Będąc bardzo małymi, mogą tworzyć się w dużych ilościach i być przenoszone przez wiatr z dala od rodzicielskiego sporofitu. Przypadkowo mogą znajdować się w pobliżu megaspory, która w roślinach nasiennych nie oddziela się od rodzicielskiego sporofitu (ryc. 2.45). Dokładnie tak to się dzieje zapylanie u roślin, których ziarna pyłku są mikrosporami. Gamety męskie są produkowane w ziarnach pyłku.

Ryż. 2.45. Schematyczne przedstawienie głównych elementów różnorodności i zapylania.
Rośliny nasienne rozwinęły jeszcze jedną ewolucyjną zaletę. Męskie gamety nie muszą już podpływać do żeńskich gamet, ponieważ rośliny nasienne wyewoluowały łagiewki pyłkowe. Rozwijają się z ziaren pyłku i rosną w kierunku gamet żeńskich. Przez tę rurkę męskie gamety docierają do żeńskiej gamety i ją zapładniają. Pływające plemniki już się nie tworzą, w zapłodnieniu biorą udział tylko męskie jądra.
W konsekwencji rośliny wykształciły mechanizm zapłodnienia niezależny od wody. To był jeden z powodów, dla których rośliny nasienne były tak lepsze od innych roślin w rozwoju ziemi. Początkowo zapylanie odbywało się tylko za pomocą wiatru - dość losowy proces, któremu towarzyszyły duże straty pyłku. Jednak już we wczesnych stadiach ewolucji, około 300 milionów lat temu w okresie karbońskim, pojawiły się owady latające, a wraz z nimi możliwość efektywniejszego zapylania. Rośliny kwitnące intensywnie wykorzystują zapylanie przez owady, podczas gdy w drzewach iglastych nadal dominuje zapylenie wiatrem.
POSIEW. We wczesnych roślinach heterosporowych megaspory były uwalniane z macierzystego sporofitu, podobnie jak mikrospory. W roślinach nasiennych megaspory nie oddzielają się od rośliny macierzystej, pozostając w megasporangii lub owule(rys. 2.45). Zalążek zawiera gametę żeńską. Po zapłodnieniu gamet żeńskich zalążki są już nazywane nasionko. Zatem nasienie jest zapłodnioną zalążkiem. Obecność zalążka i nasiona daje pewne korzyści roślinom nasiennym.
- Gametofit żeński jest chroniony przez zalążek. Jest całkowicie zależny od rodzicielskiego sporofitu iw przeciwieństwie do wolno żyjącego gametofitu jest niewrażliwy na odwodnienie.
- Po zapłodnieniu ziarno tworzy rezerwę składników odżywczych otrzymywanych przez gametofit z rodzicielskiej rośliny sporofitu, od którego nadal nie jest oddzielony. Ta rezerwa jest wykorzystywana przez rozwijającą się zygotę (kolejne pokolenie sporofitów) po kiełkowaniu nasion.
- Nasiona są zaprojektowane tak, aby przetrwać niekorzystne warunki i pozostać w stanie uśpienia, dopóki warunki nie będą sprzyjać kiełkowaniu.
- Nasiona mogą rozwijać różne adaptacje, aby ułatwić ich rozsiewanie.
Nasienie jest złożoną strukturą, w której zgromadzone są komórki trzech pokoleń - sporofit rodzicielski, gametofit żeński i zarodek kolejnego pokolenia sporofitów. Sporofit rodzicielski dostarcza nasionom wszystko, czego potrzebuje do życia, i to dopiero po pełnym dojrzeniu nasion, tj. gromadzi zapasy składników odżywczych dla zarodka sporofitu, oddziela się od rodzicielskiego sporofitu.
2.7. Szanse na przetrwanie i rozwój ziaren pyłku przenoszonego przez wiatr (mikrospory) są znacznie mniejsze niż w przypadku zarodników Dryopteris. Czemu?
2.8. Wyjaśnij, dlaczego megaspory są duże, a mikrospory małe.
2.7.7. Krótka lista adaptacji roślin nasiennych do życia na lądzie
Główne zalety roślin nasiennych nad wszystkimi innymi są następujące.
- Wytwarzanie gametofitu jest znacznie zredukowane i całkowicie zależne od sporofitu, dobrze przystosowanego do życia na lądzie, wewnątrz którego gametofit jest zawsze chroniony. W innych roślinach gametofit bardzo łatwo wysycha.
- Nawożenie następuje niezależnie od wody. Gamety męskie są nieruchome i przenoszone w ziarnach pyłku przez wiatr lub owady. Ostateczny transfer gamet męskich do gamet żeńskich następuje za pomocą łagiewki pyłkowej.
- Zapłodnione zalążki (nasiona) pozostają przez pewien czas na macierzystym sporoficie, z którego otrzymują ochronę i pożywienie, zanim zostaną wydalone.
- W wielu roślinach nasiennych obserwuje się wzrost wtórny z odkładaniem się dużych ilości drewna, które pełni funkcję podtrzymującą. Takie rośliny wyrastają na drzewa i krzewy, które mogą skutecznie konkurować o światło i inne zasoby.
Niektóre z najważniejszych trendów ewolucyjnych podsumowano na ryc. 2.33. Rośliny nasienne mają inne cechy, które są nieodłączne od roślin nie tylko z tej grupy, ale także pełnią rolę adaptacji do życia na lądzie.

Ryż. 2.33. Systematyka roślin i wybrane główne kierunki ewolucji roślin.
- Prawdziwe korzenie zapewniają pozyskiwanie wilgoci z gleby.
- Rośliny chroni przed wysychaniem naskórek z wodoszczelnym naskórkiem (lub czopem powstałym po wzroście wtórnym).
- Naskórek naziemnych części rośliny, zwłaszcza liści, jest penetrowany przez wiele maleńkich szczelin zwanych szparki przez które następuje wymiana gazowa między instalacją a atmosferą.
- Rośliny mają również wyspecjalizowane przystosowania do życia w gorących i suchych warunkach (rozdz. 19 i 20).
Zdolność adaptacji ontogenezy roślin do warunków środowiskowych jest wynikiem ich ewolucyjnego rozwoju (zmienność, dziedziczność, selekcja). W trakcie filogenezy każdego gatunku rośliny, w procesie ewolucji, wykształciły się pewne potrzeby jednostki w zakresie warunków egzystencji i przystosowania do zajmowanej przez niego niszy ekologicznej. Tolerancja na wilgoć i cień, ciepło, zimno i inne cechy ekologiczne poszczególnych gatunków roślin ukształtowały się w toku ewolucji w wyniku długotrwałego narażenia na odpowiednie warunki. Tak więc rośliny kochające ciepło i rośliny krótkiego dnia są charakterystyczne dla południowych szerokości geograficznych, mniej wymagających ciepła i roślin długiego dnia - dla północnych.
W naturze, w jednym regionie geograficznym, każdy gatunek rośliny zajmuje niszę ekologiczną odpowiadającą jego cechom biologicznym: kochający wilgoć - bliżej zbiorników wodnych, odporny na cień - pod baldachimem lasu itp. Dziedziczność roślin powstaje pod wpływem określonych warunków środowiskowych. Nie bez znaczenia są również zewnętrzne uwarunkowania ontogenezy roślin.
W większości przypadków rośliny i uprawy (nasadzenia) upraw rolnych, doświadczające działania pewnych niekorzystnych czynników, wykazują na nie odporność w wyniku adaptacji do warunków egzystencji, które rozwinęły się historycznie, co zauważył K. A. Timiryazev.
1. Podstawowe środowiska życia.
Podczas badania środowiska (siedliska roślin i zwierząt oraz działalności produkcyjnej człowieka) wyróżnia się następujące główne elementy: środowisko powietrzne; środowisko wodne (hydrosfera); fauna (zwierzęta ludzkie, domowe i dzikie, w tym ryby i ptaki); roślinność (rośliny uprawne i dziko rosnące, w tym wodne), glebę (warstwa roślinności), podglebie (górna część skorupy ziemskiej, w obrębie której możliwa jest eksploatacja); środowisko klimatyczne i akustyczne.
Środowisko powietrzne może być zewnętrzne, w którym większość ludzi spędza mniej czasu (do 10-15%), produkcja wewnętrzna (osoba spędza w nim do 25-30% czasu) i wewnętrzne mieszkalne, gdzie ludzie pozostają przez większość czasu (do 60 -70% lub więcej).
Powietrze zewnętrzne na powierzchni ziemi zawiera objętościowo: 78,08% azotu; 20,95% tlenu; 0,94% gazów obojętnych i 0,03% dwutlenku węgla. Na wysokości 5 km zawartość tlenu pozostaje taka sama, a azot wzrasta do 78,89%. Często powietrze przy powierzchni ziemi zawiera różne zanieczyszczenia, zwłaszcza w miastach: tam zawiera ponad 40 składników obcych dla naturalnego środowiska powietrza. Powietrze wewnętrzne w mieszkaniach z reguły ma
zwiększona zawartość dwutlenku węgla, a powietrze wewnętrzne pomieszczeń przemysłowych zwykle zawiera zanieczyszczenia, których charakter określa technologia produkcji. Wśród gazów uwalniana jest para wodna, która przedostaje się do atmosfery w wyniku parowania z Ziemi. Większość (90%) jest skoncentrowana w najniższej pięciokilometrowej warstwie atmosfery, z wysokością jej ilość bardzo szybko maleje. Atmosfera zawiera dużo pyłu, który dostaje się tam z powierzchni Ziemi i częściowo z kosmosu. Podczas silnych fal wiatry zbierają rozpryski wody z mórz i oceanów. W ten sposób cząsteczki soli przedostają się z wody do atmosfery. W wyniku erupcji wulkanów, pożarów lasów, obiektów przemysłowych itp. powietrze jest zanieczyszczone produktami niepełnego spalania. W przyziemnej warstwie powietrza znajduje się przede wszystkim pył i inne zanieczyszczenia. Nawet po deszczu 1 cm zawiera około 30 tysięcy cząsteczek kurzu, a przy suchej pogodzie jest ich kilkakrotnie więcej przy bezdeszczowej pogodzie.
Wszystkie te drobne zanieczyszczenia wpływają na kolor nieba. Cząsteczki gazów rozpraszają krótkofalową część widma wiązki słonecznej, tj. fioletowe i niebieskie promienie. Tak więc w ciągu dnia niebo jest niebieskie. A cząsteczki zanieczyszczeń, które są znacznie większe niż cząsteczki gazu, rozpraszają promienie świetlne o prawie wszystkich długościach fal. Dlatego gdy powietrze jest zakurzone lub zawiera kropelki wody, niebo staje się białawe. Na dużych wysokościach niebo jest ciemnofioletowe, a nawet czarne.
W wyniku zachodzącej na Ziemi fotosyntezy roślinność rocznie wytwarza 100 miliardów ton substancji organicznych (około połowy przypada na morza i oceany), asymilując około 200 miliardów ton dwutlenku węgla i uwalniając do środowiska około 145 miliardów ton . wolny tlen uważa się, że w wyniku fotosyntezy powstaje cały tlen w atmosferze. Na rolę terenów zielonych w tym cyklu wskazują następujące dane: 1 hektar terenów zielonych oczyszcza powietrze z 8 kg dwutlenku węgla średnio w ciągu 1 godziny (200 osób emitowanych w tym czasie podczas oddychania). Dorosłe drzewo uwalnia dziennie 180 litrów tlenu, aw ciągu pięciu miesięcy (od maja do września) pochłania około 44 kg dwutlenku węgla.
Ilość uwalnianego tlenu i pochłanianego dwutlenku węgla zależy od wieku terenów zielonych, składu gatunkowego, gęstości sadzenia i innych czynników.
Równie ważne są rośliny morskie – fitoplankton (głównie glony i bakterie), które w procesie fotosyntezy uwalniają tlen.
Środowisko wodne obejmuje wody powierzchniowe i podziemne. Wody powierzchniowe koncentrują się głównie w oceanach, ich zawartość to 1 miliard 375 milionów kilometrów sześciennych - około 98% wszystkich wód na Ziemi. Powierzchnia oceanu (powierzchnia wodna) wynosi 361 milionów kilometrów kwadratowych. Jest to około 2,4 razy większa powierzchnia lądu - terytorium, które zajmuje 149 milionów kilometrów kwadratowych. Woda w oceanie jest słona, a większość z niej (ponad 1 miliard kilometrów sześciennych) zachowuje stałe zasolenie około 3,5% i temperaturę około 3,7°C. Zauważalne różnice w zasoleniu i temperaturze obserwuje się prawie wyłącznie na powierzchni warstwy wody, a także w marginalnych, a zwłaszcza w morzach Śródziemnomorskich. Zawartość tlenu rozpuszczonego w wodzie znacznie spada na głębokości 50-60 metrów.
Wody gruntowe mogą być słone, słonawe (mniejsze zasolenie) i świeże; istniejące wody geotermalne mają podwyższoną temperaturę (ponad 30ºC).
Do działalności produkcyjnej ludzkości i jej potrzeb domowych potrzebna jest świeża woda, której ilość stanowi zaledwie 2,7% całkowitej objętości wody na Ziemi, a bardzo mała jej część (tylko 0,36%) jest dostępna w miejscach, które są łatwo dostępne do ekstrakcji. Większość słodkiej wody znajduje się w śniegu i słodkowodnych górach lodowych znajdujących się głównie na obszarze koła podbiegunowego.
Roczny globalny odpływ słodkiej wody rzecznej wynosi 37,3 tys. kilometrów sześciennych. Ponadto można wykorzystać część wód gruntowych równą 13 tysięcy kilometrów sześciennych. Niestety większość rzeki płynącej w Rosji, wynosząca około 5000 kilometrów sześciennych, opada na marginalne i słabo zaludnione terytoria północne.
Środowisko klimatyczne jest ważnym czynnikiem determinującym rozwój różnych gatunków flory i fauny oraz jej żyzność. Charakterystyczną cechą Rosji jest to, że większość jej terytorium ma znacznie chłodniejszy klimat niż w innych krajach.
Wszystkie rozważane komponenty środowiska są zawarte w
BIOSFERA: powłoka Ziemi, w tym część atmosfery, hydrosfera i górna część litosfery, które są połączone złożonymi cyklami biochemicznymi migracji materii i energii, powłoka geologiczna Ziemi, zamieszkana przez organizmy żywe. Górna granica życia biosfery jest ograniczona przez intensywne stężenie promieni ultrafioletowych; niższa - wysoka temperatura wnętrza ziemi (powyżej 100`C). Jego skrajne granice osiągają tylko organizmy niższe - bakterie.
Adaptację (adaptację) rośliny do określonych warunków środowiskowych zapewniają mechanizmy fizjologiczne (adaptacja fizjologiczna), aw populacji organizmów (gatunków) - mechanizmy zmienności genetycznej, dziedziczności i selekcji (adaptacja genetyczna). Czynniki środowiskowe mogą zmieniać się regularnie i losowo. Regularnie zmieniające się warunki środowiskowe (zmiana pór roku) rozwijają w roślinach adaptację genetyczną do tych warunków.
W naturalnych warunkach wzrostu lub uprawy gatunku, w trakcie swojego wzrostu i rozwoju, często doświadczają one wpływu niekorzystnych czynników środowiskowych, do których należą wahania temperatury, susza, nadmierna wilgoć, zasolenie gleby itp. Każda roślina ma umiejętność adaptacji do zmieniających się warunków środowiskowych w granicach wyznaczonych przez jego genotyp. Im wyższa zdolność rośliny do zmiany metabolizmu zgodnie ze środowiskiem, tym większa szybkość reakcji tej rośliny i lepsza zdolność adaptacji. Ta właściwość wyróżnia odporne odmiany upraw rolniczych. Z reguły niewielkie i krótkotrwałe zmiany czynników środowiskowych nie prowadzą do istotnych zaburzeń funkcji fizjologicznych roślin, co wynika z ich zdolności do utrzymywania względnie stabilnego stanu w zmieniających się warunkach środowiskowych, czyli utrzymania homeostazy. Jednak ostre i długotrwałe uderzenia prowadzą do zakłócenia wielu funkcji rośliny, a często do jej śmierci.
Pod wpływem niesprzyjających warunków obniżenie procesów i funkcji fizjologicznych może osiągnąć poziomy krytyczne, które nie zapewniają realizacji genetycznego programu ontogenezy, zaburzony zostaje metabolizm energetyczny, układy regulacyjne, metabolizm białek i inne funkcje życiowe organizmu roślinnego. Kiedy roślina jest narażona na niekorzystne czynniki (stresory), powstaje w niej stan stresu, odchylenie od normy - stres. Stres to ogólna nieswoista reakcja adaptacyjna organizmu na działanie jakichkolwiek niekorzystnych czynników. Istnieją trzy główne grupy czynników wywołujących stres u roślin: fizyczne - niedostateczna lub nadmierna wilgotność, światło, temperatura, promieniowanie radioaktywne, stres mechaniczny; chemiczne - sole, gazy, ksenobiotyki (herbicydy, insektycydy, fungicydy, odpady przemysłowe itp.); biologiczne - uszkodzenia przez patogeny lub szkodniki, konkurencja z innymi roślinami, wpływ zwierząt, kwitnienie, dojrzewanie owoców.
Siła stresu zależy od tempa rozwoju niekorzystnej dla rośliny sytuacji oraz poziomu czynnika stresowego. Przy powolnym rozwoju niesprzyjających warunków roślina lepiej się do nich przystosowuje niż przy krótkotrwałym, ale silnym działaniu. W pierwszym przypadku z reguły w większym stopniu przejawiają się specyficzne mechanizmy oporu, w drugim - niespecyficzne.
W niesprzyjających warunkach naturalnych o odporności i produktywności roślin decyduje szereg znaków, właściwości oraz reakcje ochronne i adaptacyjne. Różne gatunki roślin zapewniają stabilność i przetrwanie w niesprzyjających warunkach na trzy główne sposoby: poprzez mechanizmy, które pozwalają im uniknąć niekorzystnych skutków (uśpienie, przetrwanie itp.); poprzez specjalne urządzenia konstrukcyjne; ze względu na właściwości fizjologiczne, które pozwalają im przezwyciężyć szkodliwy wpływ środowiska.
Rośliny uprawne jednoroczne w strefach umiarkowanych, uzupełniając swoją ontogenezę w stosunkowo sprzyjających warunkach, zimują w postaci stabilnych nasion (stan spoczynku). Wiele bylin zimuje jako podziemne organy magazynujące (cebule lub kłącza) chronione przed zamarzaniem warstwą gleby i śniegu. Drzewa i krzewy owocowe ze stref umiarkowanych, chroniące się przed zimowym chłodem, zrzucają liście.
Ochronę przed niekorzystnymi czynnikami środowiskowymi w roślinach zapewniają adaptacje strukturalne, cechy budowy anatomicznej (naskórek, skorupa, tkanki mechaniczne itp.), specjalne narządy ochronne (palenie włosów, kolce), reakcje motoryczne i fizjologiczne oraz wytwarzanie ochronnych substancje (żywice, fitoncydy, toksyny, białka ochronne).
Adaptacje strukturalne obejmują liście drobnolistne, a nawet brak liści, woskowatą skórkę na powierzchni liści, ich gęste omijanie i zanurzenie szparek, obecność soczystych liści i łodyg zatrzymujących zapasy wody, erekcje lub opadające liście itp. Rośliny posiadają różne mechanizmy fizjologiczne, które pozwalają im przystosować się do niesprzyjających warunków środowiskowych. Jest to samoistny rodzaj fotosyntezy u sukulentów, minimalizujący utratę wody i niezbędny do przetrwania roślin na pustyni itp.
2. Adaptacja w roślinach
Tolerancja roślin na zimno
Odporność roślin na niskie temperatury dzieli się na mrozoodporność i mrozoodporność. Odporność na zimno rozumiana jest jako zdolność roślin do tolerowania dodatnich temperatur nieco wyższych niż 0 C. Odporność na zimno jest charakterystyczna dla roślin strefy umiarkowanej (jęczmień, owies, len, wyka itp.). Rośliny tropikalne i subtropikalne ulegają uszkodzeniu i obumierają w temperaturze od 0º do 10ºC (kawa, bawełna, ogórek itp.). Dla większości roślin rolniczych niskie temperatury dodatnie nie są szkodliwe. Wynika to z faktu, że podczas chłodzenia aparat enzymatyczny roślin nie jest zaburzony, odporność na choroby grzybowe nie zmniejsza się i w ogóle nie dochodzi do zauważalnych uszkodzeń roślin.
Stopień odporności na zimno różnych roślin nie jest taki sam. Wiele roślin na południowych szerokościach geograficznych jest niszczonych przez zimno. W temperaturze 3 ° C uszkadza się ogórek, bawełna, fasola, kukurydza i bakłażan. Odmiany różnią się tolerancją na zimno. Do scharakteryzowania odporności roślin na zimno stosuje się pojęcie minimalnej temperatury, przy której zatrzymuje się wzrost roślin. Dla dużej grupy roślin rolniczych jego wartość wynosi 4°C. Jednak wiele roślin ma wyższą temperaturę minimalną i dlatego jest mniej odporne na zimno.
Adaptacja roślin do niskich temperatur dodatnich.
Odporność na niskie temperatury jest cechą uwarunkowaną genetycznie. Odporność roślin na zimno zależy od zdolności roślin do utrzymania prawidłowej struktury cytoplazmy, zmiany metabolizmu w okresie schładzania i późniejszego wzrostu temperatury na wystarczająco wysokim poziomie.
Mrozoodporność roślin
Mrozoodporność - zdolność roślin do tolerowania temperatur poniżej 0 ° C, niskich ujemnych temperatur. Rośliny odporne na mróz są w stanie zapobiegać lub redukować wpływ niskich ujemnych temperatur. Zimą przymrozki o temperaturze poniżej -20 ° C są powszechne na znacznej części terytorium Rosji. Na mróz narażone są rośliny jednoroczne, dwuletnie i wieloletnie. Rośliny znoszą warunki zimowe w różnych okresach rozwoju osobniczego. W uprawach jednorocznych nasiona (rośliny jare), rośliny porośnięte (rośliny ozime) zimują, w uprawach dwuletnich i wieloletnich - bulwy, rośliny okopowe, cebulki, kłącza, rośliny dorosłe. Zdolność zimowych, wieloletnich roślin zielnych i drzewiastych do zimowania wynika z ich dość wysokiej mrozoodporności. Tkanki tych roślin mogą zamarznąć, ale rośliny nie obumierają.
Zamrażanie komórek i tkanek roślinnych oraz zachodzące podczas tego procesy.
O zdolności roślin do tolerowania ujemnych temperatur decyduje podstawa dziedziczna danego gatunku rośliny, jednak mrozoodporność jednej i tej samej rośliny zależy od warunków poprzedzających nadejście mrozu, wpływających na charakter zlodzenia. Lód może tworzyć się zarówno w protoplastach komórki, jak iw przestrzeni międzykomórkowej. Nie każda formacja lodu powoduje obumieranie komórek roślinnych.
Stopniowy spadek temperatury w tempie 0,5–1°C/h prowadzi do powstawania kryształków lodu, głównie w przestrzeniach międzykomórkowych i początkowo nie powoduje śmierci komórki. Jednak konsekwencje tego procesu mogą być szkodliwe dla komórki. Tworzenie się lodu w protoplastach komórki z reguły następuje z szybkim spadkiem temperatury. Następuje koagulacja białek protoplazmatycznych, struktury komórkowe są uszkadzane przez kryształki lodu powstające w cytozolu, komórki umierają. Rośliny zabite przez mróz po rozmrożeniu tracą turgor, woda wypływa z ich mięsistych tkanek.
Rośliny odporne na mróz mają adaptacje, które zmniejszają odwodnienie komórek. Wraz ze spadkiem temperatury rośliny takie wykazują wzrost zawartości cukrów i innych substancji chroniących tkanki (krioprotektory), są to przede wszystkim białka hydrofilowe, mono- i oligosacharydy; spadek nawodnienia komórek; wzrost ilości polarnych lipidów i zmniejszenie nasycenia ich reszt kwasów tłuszczowych; wzrost liczby białek ochronnych.
Na stopień mrozoodporności roślin duży wpływ mają cukry, regulatory wzrostu i inne substancje powstające w komórkach. W roślinach zimujących cukry gromadzą się w cytoplazmie, a zawartość skrobi spada. Wpływ cukrów na zwiększenie mrozoodporności roślin jest wieloaspektowy. Kumulacja cukrów zapobiega zamarzaniu dużej ilości wody wewnątrzkomórkowej, znacznie zmniejsza ilość powstającego lodu.
Właściwość mrozoodporności powstaje w procesie ontogenezy roślin pod wpływem określonych warunków środowiskowych zgodnie z genotypem rośliny, związanym z gwałtownym spadkiem tempa wzrostu, przejściem rośliny w stan uśpienia.
Cykl życiowy rozwoju roślin zimowych, dwuletnich i wieloletnich jest kontrolowany przez sezonowy rytm okresów światła i temperatury. W przeciwieństwie do jednorocznych wiosennych, zaczynają przygotowywać się do znoszenia niekorzystnych warunków zimowych od momentu zaprzestania wzrostu, a następnie jesienią, gdy spadają temperatury.
Zimotrwalosc roślin
Zimotrwalosc jako odporność na kompleks niekorzystnych czynników zimujących.
Bezpośredni wpływ mrozu na komórki to nie jedyne niebezpieczeństwo, które zagraża wieloletnim uprawom zielnym i drzewiastym, roślinom ozimym w okresie zimowym. Oprócz bezpośredniego działania mrozu rośliny narażone są na szereg innych niekorzystnych czynników. W zimie temperatury mogą się znacznie wahać. Przymrozki są często zastępowane krótkotrwałymi i długotrwałymi roztopami. Zimą burze śnieżne nie są rzadkością, a w bezśnieżne zimy w bardziej południowych regionach kraju występują również suche wiatry. Wszystko to wyczerpuje rośliny, które po przezimowaniu wychodzą bardzo osłabione i mogą następnie obumrzeć.
Szczególnie liczne niekorzystne skutki odczuwają rośliny zielne wieloletnie i jednoroczne. Na terytorium Rosji w niesprzyjających latach śmierć zbóż ozimych sięga 30-60%. Giną nie tylko rośliny ozime, ale także wieloletnie trawy, plantacje owoców i jagód. Zimą i wczesną wiosną rośliny zimowe są uszkadzane i giną z powodu wielu innych niekorzystnych czynników: zwilżania, zawilgocenia, zaspy lodowej, wybrzuszeń, uszkodzeń spowodowanych zimową suszą.
Zmoczenie, przemoczenie, śmierć pod lodową skorupą, wybrzuszenie, zimowa susza.
Tłumienie. Wśród wymienionych przeciwności pierwsze miejsce zajmuje rozkład roślin. Zamieranie roślin na skutek zamrożenia obserwuje się głównie w ciepłe zimy z dużą pokrywą śnieżną, która utrzymuje się przez 2-3 miesiące, zwłaszcza gdy śnieg pada na mokry i rozmarznięty grunt. Badania wykazały, że przyczyną obumierania upraw ozimych w wyniku zamierania roślin jest zubożenie roślin. Przebywając pod śniegiem w temperaturze około 0 ° C w bardzo wilgotnym środowisku, prawie zupełnej ciemności, tj. w warunkach, w których proces oddychania jest dość intensywny i fotosynteza jest wykluczona, rośliny stopniowo zużywają cukier i inne zapasy składników odżywczych zgromadzone w tym okresie przechodzą przez pierwszą fazę twardnienia i giną z wycieńczenia (zawartość cukrów w tkankach spada z 20 do 2-4%) i przymrozków wiosennych. Wiosną takie rośliny są łatwo uszkadzane przez pleśń śnieżną, co również prowadzi do ich śmierci.
Zwilżanie. Zwilżanie występuje głównie wiosną w niskich miejscach w okresie topnienia śniegu, rzadziej podczas przedłużających się roztopów, kiedy to na powierzchni gleby gromadzi się woda roztopowa, która nie jest wchłaniana do zamarzniętej gleby i może zalewać rośliny. W tym przypadku przyczyną śmierci roślin jest ostry brak tlenu (warunki beztlenowe - niedotlenienie). W roślinach znajdujących się pod warstwą wody normalne oddychanie ustaje z powodu braku tlenu w wodzie i glebie. Brak tlenu wzmaga oddychanie beztlenowe roślin, w wyniku czego mogą powstawać substancje toksyczne i rośliny umierają z wycieńczenia i bezpośredniego zatrucia organizmu.
Śmierć pod lodową skorupą. Na polach w miejscach, gdzie częste odwilże zastępują silne mrozy, tworzy się skorupa lodowa. Efekt moczenia w tym przypadku może ulec pogorszeniu. W takim przypadku dochodzi do powstawania wiszących lub naziemnych (kontaktowych) skorup lodowych. Wiszące skorupy są mniej niebezpieczne, ponieważ tworzą się na wierzchu gleby i praktycznie nie mają kontaktu z roślinami; łatwo je zniszczyć wałkiem.
Gdy tworzy się ciągła skorupa kontaktowa z lodem, rośliny całkowicie zamarzają w lodzie, co prowadzi do ich śmierci, ponieważ rośliny już osłabione z moczenia są poddawane bardzo silnemu naciskowi mechanicznemu.
Wypukły. Uszkodzenia i obumieranie roślin w wyniku wybrzuszenia są determinowane przez pęknięcia w systemie korzeniowym. Wybrzuszenie roślin obserwuje się, gdy jesienią występują przymrozki przy braku pokrywy śnieżnej lub gdy w powierzchniowej warstwie gleby jest mało wody (podczas jesiennej suszy), a także podczas roztopów, gdy woda śnieżna ma czas na wchłonięcie gleba. W takich przypadkach zamarzanie wody nie zaczyna się od powierzchni gleby, ale na pewnej głębokości (gdzie występuje wilgoć). Warstwa lodu utworzona na głębokości stopniowo gęstnieje w wyniku ciągłego przepływu wody przez kapilary glebowe i unosi (wybrzusza) górne warstwy gleby wraz z roślinami, co prowadzi do łamania korzeni roślin, które mają przeniknął na znaczną głębokość.
Szkody spowodowane suszą zimową. Stabilna pokrywa śnieżna chroni zboża ozime przed wysychaniem zimą. Jednak w warunkach bezśnieżnej lub mało śnieżnej zimy, jak drzewa owocowe i krzewy, w wielu regionach Rosji często grozi im nadmierne wysuszenie przez stałe i silne wiatry, zwłaszcza pod koniec zimy ze znacznym ogrzewaniem przez słońce. Faktem jest, że bilans wodny roślin rozwija się wyjątkowo niekorzystnie zimą, ponieważ przepływ wody z zamarzniętej gleby praktycznie ustaje.
Aby ograniczyć parowanie wody i niekorzystne skutki zimowej suszy, gatunki drzew owocowych tworzą grubą warstwę korka na gałęziach i zrzucają liście na zimę.
Wernalizacja
Fotoperiodyczne reakcje na sezonowe zmiany długości dnia są ważne dla częstotliwości kwitnienia wielu gatunków zarówno w regionach o klimacie umiarkowanym, jak i tropikalnym. Należy jednak zauważyć, że wśród gatunków z umiarkowanych szerokości geograficznych, które wykazują reakcje fotoperiodyczne, jest stosunkowo niewiele gatunków kwitnących wiosną, chociaż stale spotykamy znaczną liczbę „kwiatów kwitnących wiosną”, a wiele z tych form kwitnących wiosną na przykład Ficariaverna, pierwiosnek (Primulavutgaris), fiołki (gatunek z rodzaju Viola) itp. wykazują wyraźne zachowanie sezonowe, pozostając wegetatywnie przez pozostałą część roku po obfitym wiosennym kwitnieniu. Można przypuszczać, że wiosenne kwitnienie jest reakcją na krótkie dni w zimie, ale w przypadku wielu gatunków wydaje się, że tak nie jest.
Oczywiście długość dnia to nie jedyny czynnik zewnętrzny, który zmienia się w ciągu roku. Oczywiste jest, że temperatura również wykazuje wyraźne wahania sezonowe, zwłaszcza w regionach o klimacie umiarkowanym, chociaż czynnik ten wykazuje znaczne wahania zarówno w ciągu dnia, jak i roku. Wiemy, że sezonowe zmiany temperatury, a także zmiany długości dnia mają istotny wpływ na kwitnienie wielu gatunków roślin.
Rodzaje roślin wymagających chłodzenia, aby przejść do kwitnienia.
Stwierdzono, że wiele gatunków, w tym rośliny jednoroczne, a także dwuletnie i wieloletnie, wymaga schłodzenia, zanim zakwitną.
Zimowe rośliny jednoroczne i dwuletnie są roślinami jednokarpowymi, które wymagają jarowizacji - pozostają wegetatywne w pierwszym sezonie wegetacyjnym i kwitną następną wiosną lub wczesnym latem w odpowiedzi na okres ochłodzenia uzyskany zimą. Potrzebę chłodzenia roślin dwuletnich w celu wywołania kwitnienia wykazano doświadczalnie w przypadku wielu gatunków, takich jak burak (Betavulgaris), seler (Apiutngraveolens), kapusta i inne odmiany uprawne z rodzaju Brassica, dzwonek (Campanulamedium), trawa księżycowa (Lunariabiennis). , naparstnica (Digitalispurpurea) i inne. Jeśli rośliny naparstnicy, które w normalnych warunkach zachowują się jak rośliny dwuletnie, czyli kwitną w drugim roku po wykiełkowaniu, są trzymane w szklarni, mogą pozostać wegetatywne przez kilka lat. Na obszarach o łagodnych zimach jarmuż może rosnąć na zewnątrz przez kilka lat bez „grotu strzały” (tj. kwitnienia) wiosną, co zwykle występuje na obszarach o mroźnych zimach. Gatunki takie koniecznie wymagają jarowizacji, ale w wielu innych gatunkach kwitnienie jest przyspieszone pod wpływem zimna, ale może również zachodzić bez jarowizacji; takie gatunki wykazujące fakultatywną potrzebę przeziębienia obejmują sałatę (Lactucasaiiva), szpinak (Spinacia oleracea) i późno kwitnący groch (Pistimsa-tivum).
Podobnie jak rośliny dwuletnie, wiele bylin wymaga ekspozycji na zimno i nie zakwitnie bez corocznego zimowego chłodu. Z pospolitych bylin pierwiosnek (Primulavulgaris), fiołki (Violaspp.), lacfiol (Cheiranthuscheirii i C. allionii), lewka (Mathiolaincarna), niektóre odmiany chryzantem (Chrisant-hemummorifolium), gatunki z rodzaju Aster, goździk turecki ( Dianthus ), plewy (Loliumperenne). Gatunki wieloletnie wymagają renalizacji każdej zimy.
Prawdopodobnie można znaleźć inne kwitnące wiosną byliny, które wymagają chłodzenia. Rośliny cebulowe kwitnące na wiosnę, takie jak żonkile, hiacynty, jagody (Endymionnonscriptus), krokusy itp., nie wymagają chłodzenia w celu rozpoczęcia kwitnienia, ponieważ zawiązki kwiatów zostały zasiedlone w cebulce poprzedniego lata, ale ich wzrost jest silnie uzależniony od warunków temperaturowych . Np. u tulipana początku kwitnienia sprzyjają stosunkowo wysokie temperatury (20°C), ale dla wydłużenia łodygi i wzrostu liści optymalna temperatura początkowo wynosi 8-9°C, a w późniejszych stadiach stopniowy wzrost do 13, 17 i 23°C. Podobne reakcje na temperaturę charakteryzują hiacynty i żonkile.
U wielu gatunków inicjacja kwitnienia nie następuje podczas samego okresu schładzania i rozpoczyna się dopiero po wystawieniu rośliny na działanie wyższych temperatur po ochłodzeniu.
Tak więc, chociaż metabolizm większości roślin znacznie spowalnia w niskich temperaturach, nie ulega wątpliwości, że wernalizacja obejmuje aktywne procesy fizjologiczne, których natura jest jeszcze zupełnie nieznana.
Odporność na ciepło roślin
Odporność na ciepło (tolerancja ciepła) - zdolność roślin do wytrzymywania działania wysokich temperatur, przegrzania. Jest to cecha uwarunkowana genetycznie. Gatunki roślin różnią się tolerancją na wysokie temperatury.
Według odporności na ciepło rozróżnia się trzy grupy roślin.
Żaroodporne - ciepłolubne sinice i bakterie z gorących źródeł mineralnych, wytrzymujące temperatury do 75-100°C. O odporności cieplnej mikroorganizmów termofilnych decyduje wysoki poziom metabolizmu, zwiększona zawartość RNA w komórkach oraz odporność białka cytoplazmatycznego na koagulację termiczną.
Odporny na ciepło - rośliny pustynnych i suchych siedlisk (sukulenty, niektóre kaktusy, członkowie rodziny Grubosz), wytrzymujące nagrzewanie światłem słonecznym do 50-65ºС. Odporność na ciepło sukulentów jest w dużej mierze zdeterminowana przez zwiększoną lepkość cytoplazmy i zawartość związanej wody w komórkach oraz zmniejszony metabolizm.
Nieodporny na ciepło - rośliny mezofityczne i wodne. Mezofity miejsc otwartych tolerują krótkotrwałe narażenie na temperatury 40-47 °C, miejsca zacienione - ok. 40-42 °C, rośliny wodne wytrzymują temperatury do 38-42 °C. Spośród upraw rolnych najbardziej odporne na ciepło są rośliny ciepłolubne z południowych szerokości geograficznych (sorgo, ryż, bawełna, rącznik pospolity itp.).
Wiele mezofitów toleruje wysokie temperatury powietrza i unika przegrzania z powodu intensywnej transpiracji, która obniża temperaturę liści. Bardziej żaroodporne mezofity wyróżniają się zwiększoną lepkością cytoplazmy i zwiększoną syntezą żaroodpornych białek enzymatycznych.
Rośliny rozwinęły system adaptacji morfologicznych i fizjologicznych, które chronią je przed uszkodzeniami termicznymi: jasny kolor powierzchni, który odzwierciedla nasłonecznienie; składanie i skręcanie liści; pokwitanie lub łuski, które chronią głębsze tkanki przed przegrzaniem; cienkie warstwy tkanki korkowej, które chronią łyko i kambium; większa grubość warstwy naskórka; wysoka zawartość węglowodanów i niska - woda w cytoplazmie itp.
Rośliny bardzo szybko reagują na stres cieplny poprzez adaptację indukcyjną. Na działanie wysokich temperatur mogą przygotować się w ciągu kilku godzin. Tak więc w upalne dni odporność roślin na wysokie temperatury po południu jest wyższa niż rano. Zwykle ta stabilność jest tymczasowa, nie konsoliduje się i dość szybko znika, jeśli się ochłodzi. Odwracalność ekspozycji termicznej może wynosić od kilku godzin do 20 dni. Podczas powstawania narządów generatywnych zmniejsza się odporność cieplna roślin jednorocznych i dwuletnich.
Tolerancja roślin na suszę
Susze stały się częstym zjawiskiem w wielu regionach Rosji i krajów WNP. Susza to długi okres bezdeszczowy, któremu towarzyszy spadek względnej wilgotności powietrza, wilgotności gleby i wzrost temperatury, kiedy nie są zaspokajane normalne potrzeby wodne roślin. Na terytorium Rosji występują regiony o niestabilnej wilgotności z rocznymi opadami 250-500 mm i regiony suche, z opadami mniejszymi niż 250 mm rocznie i szybkością parowania powyżej 1000 mm.
Odporność na suszę – zdolność roślin do wytrzymywania długich okresów suszy, znaczny niedobór wody, odwodnienie komórek, tkanek i narządów. Jednocześnie szkody w uprawie zależą od czasu trwania suszy i jej intensywności. Rozróżnij suszę glebową od suszy atmosferycznej.
Susza glebowa spowodowana jest długotrwałym brakiem opadów w połączeniu z wysoką temperaturą powietrza i nasłonecznieniem, zwiększonym parowaniem z powierzchni gleby i transpiracją oraz silnymi wiatrami. Wszystko to prowadzi do przesuszenia warstwy korzeniowej gleby, zmniejszenia zaopatrzenia roślin w wodę przy niskiej wilgotności powietrza. Susza atmosferyczna charakteryzuje się wysoką temperaturą i niską wilgotnością względną (10-20%). Ciężka susza atmosferyczna jest wywoływana przez ruch mas suchego i gorącego powietrza - suchy wiatr. Zamglenie prowadzi do poważnych konsekwencji, gdy suchemu wiatrowi towarzyszy pojawienie się cząstek gleby w powietrzu (burze piaskowe).
Susza atmosferyczna, gwałtownie zwiększająca parowanie wody z powierzchni gleby i transpirację, przyczynia się do zakłócenia konsystencji tempa przedostawania się wody z gleby do organów nadziemnych i jej utraty przez roślinę, w wyniku czego roślina więdnie . Jednak przy dobrym rozwoju systemu korzeniowego susza atmosferyczna nie powoduje większych szkód dla roślin, jeśli temperatura nie przekracza granicy tolerowanej przez rośliny. Przedłużająca się susza atmosferyczna przy braku opadów prowadzi do suszy glebowej, która jest bardziej niebezpieczna dla roślin.
Odporność na suszę wynika z uwarunkowanej genetycznie zdolności przystosowania się roślin do warunków siedliskowych, a także przystosowania się do braku wody. Odporność na suszę wyraża się w zdolności roślin do znoszenia znacznego odwodnienia ze względu na rozwój wysokiego potencjału wodnego tkanek z funkcjonalnym zachowaniem struktur komórkowych, a także ze względu na adaptacyjne cechy morfologiczne łodygi, liści, organów generatywnych, które zwiększają ich wytrzymałość, tolerancję na skutki długotrwałej suszy.
Rodzaje roślin a reżim wodny
Rośliny z suchych regionów nazywane są kserofity (od greckiego xeros - suche). Są w stanie w procesie rozwoju osobniczego przystosować się do suszy atmosferycznej i glebowej. Cechą charakterystyczną kserofitów jest niewielka wielkość ich powierzchni parowania, a także niewielka wielkość części nadziemnej w porównaniu z podziemną. Kserofity to zazwyczaj zioła lub skarłowaciałe krzewy. Są podzielone na kilka typów. Przedstawiamy klasyfikację kserofitów wg P. A. Genkela.
Sukulenty są bardzo odporne na przegrzanie i odporne na odwodnienie, podczas suszy nie brakuje im wody, ponieważ zawierają jej dużą ilość i powoli ją konsumują. Ich system korzeniowy jest rozgałęziony we wszystkich kierunkach w górnych warstwach gleby, dzięki czemu rośliny szybko wchłaniają wodę w okresach deszczowych. Są to kaktusy, aloes, rozchodnik, młode.
Eukserofity to rośliny odporne na ciepło, które dobrze tolerują suszę. Do tej grupy należą rośliny stepowe, takie jak Veronica szara, aster włochaty, piołun niebieski, kolocynta arbuza, cierń wielbłąda itp. Mają niską transpirację, wysokie ciśnienie osmotyczne, cytoplazma jest wysoce elastyczna i lepka, system korzeniowy jest bardzo rozgałęziony, a jego masę umieszcza się w górnej warstwie gleby (50-60 cm). Te kserofity są zdolne do zrzucania liści, a nawet całych gałęzi.
Hemikserofity lub półkserofity to rośliny, które nie tolerują odwodnienia i przegrzania. Lepkość i elastyczność ich protoplastu jest nieznaczna, charakteryzuje się wysoką transpiracją, głębokim systemem korzeniowym, który może dotrzeć do wody podziemnej, co zapewnia nieprzerwane dostarczanie wody do rośliny. Ta grupa obejmuje szałwię, pospolitą kutrę itp.
Stipakserofshpy to trawa pierzasta, tyrsa i inne wąskolistne trawy stepowe. Są odporne na przegrzanie, dobrze wykorzystują wilgoć z krótkotrwałych deszczy. Wytrzymują tylko krótkotrwały brak wody w glebie.
Poikilokserofity to rośliny, które nie regulują swojego reżimu wodnego. Są to głównie porosty, które mogą wysychać do stanu powietrznie suchego i ponownie uaktywniać się po opadach deszczu.
Hygrofity (z greckiego hihros - mokre). Rośliny należące do tej grupy nie posiadają adaptacji ograniczających zużycie wody. Higrofity charakteryzują się stosunkowo dużymi rozmiarami komórek, cienkościenną skorupą, słabo zdrewniałymi ścianami naczyń, włóknami drzewnymi i łykowymi, cienką kutykulą i lekko pogrubionymi ściankami zewnętrznymi naskórka, dużymi aparatami szparkowymi i niewielką ich liczbą na jednostkę powierzchni, duża blaszka liściowa, słabo rozwinięta tkanka mechaniczna, rzadka sieć nerwów w liściu, duża transpiracja kutykularna, długa łodyga, słabo rozwinięty system korzeniowy. Ze względu na strukturę higrofity zbliżają się do roślin tolerujących cień, ale mają osobliwą strukturę higromorficzną. Niewielki brak wody w glebie powoduje szybkie więdnięcie higrofitów. Ciśnienie osmotyczne soku komórkowego w nich jest niskie. Należą do nich mannik, dziki rozmaryn, żurawina, przyssawka.
W zależności od warunków wzrostu i cech strukturalnych rośliny o liściach częściowo lub całkowicie zanurzonych w wodzie lub unoszących się na jej powierzchni, zwane hydrofitami, są bardzo zbliżone do higrofitów.
Mezofity (z greckiego mesos - średnie, pośrednie). Rośliny z tej grupy ekologicznej rosną w warunkach dostatecznej wilgotności. Ciśnienie osmotyczne soku komórkowego w mezofitach wynosi 1-1,5 tys. kPa. Łatwo więdną. Do mezofitów zalicza się większość traw łąkowych i strączkowych - perz rozłogowy, wyczyniec łąkowy, tymotka łąkowa, lucerna błękitna itp. Z upraw polowych, pszenica twarda i miękka, kukurydza, owies, groch, soja, burak cukrowy, konopie, prawie wszystkie owoce (z z wyjątkiem migdałów, winogron), wielu roślin warzywnych (marchew, pomidory itp.).
Narządy transpirujące - liście charakteryzują się znaczną plastycznością; w zależności od warunków wzrostu w ich strukturze obserwuje się dość duże różnice. Nawet liście tej samej rośliny o różnym zaopatrzeniu w wodę i oświetleniu mają różną strukturę. W strukturze liści ustalono pewne wzory, w zależności od ich umiejscowienia na roślinie.
V. R. Zalensky odkrył zmiany w budowie anatomicznej liści według poziomów. Odkrył, że liście górnej warstwy wykazują regularne zmiany w kierunku zwiększonego kseromorfizmu, tj. tworzą się struktury zwiększające odporność tych liści na suszę. Liście znajdujące się w górnej części łodygi zawsze różnią się od dolnych, mianowicie: im wyżej liść znajduje się na łodydze, tym mniejsze są jego komórki, tym większa liczba aparatów szparkowych i im mniejsza ich wielkość, im większa liczba włosów na jednostkę powierzchni, im gęstsza sieć wiązek naczyniowych, tym silniejsza tkanka palisadowa. Wszystkie te znaki charakteryzują kserofilię, czyli tworzenie struktur, które przyczyniają się do wzrostu odporności na suszę.
Z cechami fizjologicznymi wiąże się również pewna budowa anatomiczna, a mianowicie: górne liście wyróżniają się wyższą zdolnością asymilacyjną i intensywniejszą transpiracją. Stężenie soku w górnych liściach jest również wyższe i dlatego woda może być odciągana z dolnych liści przez górne liście, wysuszając i zamierając dolne liście. Strukturę narządów i tkanek, która zwiększa odporność roślin na suszę, nazywamy kseromorfizmem. Charakterystyczne cechy struktury liści górnej warstwy tłumaczy się tym, że rozwijają się one w warunkach nieco trudnego zaopatrzenia w wodę.
Powstał złożony system adaptacji anatomicznych i fizjologicznych, aby wyrównać równowagę między dopływem i odpływem wody w roślinie. Takie adaptacje obserwuje się u kserofitów, higrofitów, mezofitów.
Wyniki badań wykazały, że właściwości adaptacyjne form roślin odpornych na suszę powstają pod wpływem warunków ich istnienia.
WNIOSEK
Niesamowitą harmonię żywej natury, jej doskonałość tworzy sama natura: walka o przetrwanie. Formy adaptacji roślin i zwierząt są nieskończenie różnorodne. Od czasu swojego pojawienia się cały świat zwierząt i roślin doskonalił się na drodze celowych adaptacji do warunków życia: do wody, powietrza, światła słonecznego, grawitacji itp.
LITERATURA
1. Wołodko I.K. „Mikroelementy i odporność roślin na niekorzystne warunki”, Mińsk, Nauka i technologia, 1983.
2. Goryszyna T.K. „„Ekologia roślin””, uch. Podręcznik dla uniwersytetów, Moskwa, V. school, 1979.
3. Prokofiew A.A. „Problemy odporności roślin na suszę”, Moskwa, Nauka, 1978.
4. Sergeeva K.A. „„ Fizjologiczne i biochemiczne podstawy zimotrwałości roślin drzewiastych „”, Moskwa, Nauka, 1971
5. Kultiasow I.M. Ekologia roślin. - M.: Wydawnictwo Uniwersytetu Moskiewskiego, 1982
Zdolność adaptacji ontogenezy roślin do warunków środowiskowych jest wynikiem ich ewolucyjnego rozwoju (zmienność, dziedziczność, selekcja). W trakcie filogenezy każdego gatunku rośliny, w procesie ewolucji, wykształciły się pewne potrzeby jednostki w zakresie warunków egzystencji i przystosowania do zajmowanej przez niego niszy ekologicznej. Tolerancja na wilgoć i cień, ciepło, zimno i inne cechy ekologiczne poszczególnych gatunków roślin ukształtowały się w toku ewolucji w wyniku długotrwałego narażenia na odpowiednie warunki. Tak więc rośliny kochające ciepło i rośliny krótkiego dnia są charakterystyczne dla południowych szerokości geograficznych, mniej wymagających ciepła i roślin długiego dnia - dla północnych.
W naturze, w jednym regionie geograficznym, każdy gatunek rośliny zajmuje niszę ekologiczną odpowiadającą jego cechom biologicznym: kochający wilgoć - bliżej zbiorników wodnych, odporny na cień - pod baldachimem lasu itp. Dziedziczność roślin powstaje pod wpływem określonych warunków środowiskowych. Nie bez znaczenia są również zewnętrzne uwarunkowania ontogenezy roślin.
W większości przypadków rośliny i uprawy (nasadzenia) upraw rolnych, doświadczające działania pewnych niekorzystnych czynników, wykazują na nie odporność w wyniku adaptacji do warunków egzystencji, które rozwinęły się historycznie, co zauważył K. A. Timiryazev.
1. Podstawowe środowiska życia.
Podczas badania środowiska (siedliska roślin i zwierząt oraz działalności produkcyjnej człowieka) wyróżnia się następujące główne elementy: środowisko powietrzne; środowisko wodne (hydrosfera); fauna (zwierzęta ludzkie, domowe i dzikie, w tym ryby i ptaki); roślinność (rośliny uprawne i dziko rosnące, w tym wodne), glebę (warstwa roślinności), podglebie (górna część skorupy ziemskiej, w obrębie której możliwa jest eksploatacja); środowisko klimatyczne i akustyczne.
Środowisko powietrzne może być zewnętrzne, w którym większość ludzi spędza mniej czasu (do 10-15%), produkcja wewnętrzna (osoba spędza w nim do 25-30% czasu) i wewnętrzne mieszkalne, gdzie ludzie pozostają przez większość czasu (do 60 -70% lub więcej).
Powietrze zewnętrzne na powierzchni ziemi zawiera objętościowo: 78,08% azotu; 20,95% tlenu; 0,94% gazów obojętnych i 0,03% dwutlenku węgla. Na wysokości 5 km zawartość tlenu pozostaje taka sama, a azot wzrasta do 78,89%. Często powietrze przy powierzchni ziemi zawiera różne zanieczyszczenia, zwłaszcza w miastach: tam zawiera ponad 40 składników obcych dla naturalnego środowiska powietrza. Powietrze wewnętrzne w mieszkaniach z reguły ma
zwiększona zawartość dwutlenku węgla, a powietrze wewnętrzne pomieszczeń przemysłowych zwykle zawiera zanieczyszczenia, których charakter określa technologia produkcji. Wśród gazów uwalniana jest para wodna, która przedostaje się do atmosfery w wyniku parowania z Ziemi. Większość (90%) jest skoncentrowana w najniższej pięciokilometrowej warstwie atmosfery, z wysokością jej ilość bardzo szybko maleje. Atmosfera zawiera dużo pyłu, który dostaje się tam z powierzchni Ziemi i częściowo z kosmosu. Podczas silnych fal wiatry zbierają rozpryski wody z mórz i oceanów. W ten sposób cząsteczki soli przedostają się z wody do atmosfery. W wyniku erupcji wulkanów, pożarów lasów, obiektów przemysłowych itp. powietrze jest zanieczyszczone produktami niepełnego spalania. W przyziemnej warstwie powietrza znajduje się przede wszystkim pył i inne zanieczyszczenia. Nawet po deszczu 1 cm zawiera około 30 tysięcy cząsteczek kurzu, a przy suchej pogodzie jest ich kilkakrotnie więcej przy bezdeszczowej pogodzie.
Wszystkie te drobne zanieczyszczenia wpływają na kolor nieba. Cząsteczki gazów rozpraszają krótkofalową część widma wiązki słonecznej, tj. fioletowe i niebieskie promienie. Tak więc w ciągu dnia niebo jest niebieskie. A cząsteczki zanieczyszczeń, które są znacznie większe niż cząsteczki gazu, rozpraszają promienie świetlne o prawie wszystkich długościach fal. Dlatego gdy powietrze jest zakurzone lub zawiera kropelki wody, niebo staje się białawe. Na dużych wysokościach niebo jest ciemnofioletowe, a nawet czarne.
W wyniku zachodzącej na Ziemi fotosyntezy roślinność rocznie wytwarza 100 miliardów ton substancji organicznych (około połowy przypada na morza i oceany), asymilując około 200 miliardów ton dwutlenku węgla i uwalniając do środowiska około 145 miliardów ton . wolny tlen uważa się, że w wyniku fotosyntezy powstaje cały tlen w atmosferze. Na rolę terenów zielonych w tym cyklu wskazują następujące dane: 1 hektar terenów zielonych oczyszcza powietrze z 8 kg dwutlenku węgla średnio w ciągu 1 godziny (200 osób emitowanych w tym czasie podczas oddychania). Dorosłe drzewo uwalnia dziennie 180 litrów tlenu, aw ciągu pięciu miesięcy (od maja do września) pochłania około 44 kg dwutlenku węgla.
Ilość uwalnianego tlenu i pochłanianego dwutlenku węgla zależy od wieku terenów zielonych, składu gatunkowego, gęstości sadzenia i innych czynników.
Równie ważne są rośliny morskie – fitoplankton (głównie glony i bakterie), które w procesie fotosyntezy uwalniają tlen.
Środowisko wodne obejmuje wody powierzchniowe i podziemne. Wody powierzchniowe koncentrują się głównie w oceanach, ich zawartość to 1 miliard 375 milionów kilometrów sześciennych - około 98% wszystkich wód na Ziemi. Powierzchnia oceanu (powierzchnia wodna) wynosi 361 milionów kilometrów kwadratowych. Jest to około 2,4 razy większa powierzchnia lądu - terytorium, które zajmuje 149 milionów kilometrów kwadratowych. Woda w oceanie jest słona, a większość z niej (ponad 1 miliard kilometrów sześciennych) zachowuje stałe zasolenie około 3,5% i temperaturę około 3,7°C. Zauważalne różnice w zasoleniu i temperaturze obserwuje się prawie wyłącznie na powierzchni warstwy wody, a także w marginalnych, a zwłaszcza w morzach Śródziemnomorskich. Zawartość tlenu rozpuszczonego w wodzie znacznie spada na głębokości 50-60 metrów.
Wody gruntowe mogą być słone, słonawe (mniejsze zasolenie) i świeże; istniejące wody geotermalne mają podwyższoną temperaturę (ponad 30ºC).
Do działalności produkcyjnej ludzkości i jej potrzeb domowych potrzebna jest świeża woda, której ilość stanowi zaledwie 2,7% całkowitej objętości wody na Ziemi, a bardzo mała jej część (tylko 0,36%) jest dostępna w miejscach, które są łatwo dostępne do ekstrakcji. Większość słodkiej wody znajduje się w śniegu i słodkowodnych górach lodowych znajdujących się głównie na obszarze koła podbiegunowego.
Roczny globalny odpływ słodkiej wody rzecznej wynosi 37,3 tys. kilometrów sześciennych. Ponadto można wykorzystać część wód gruntowych równą 13 tysięcy kilometrów sześciennych. Niestety większość rzeki płynącej w Rosji, wynosząca około 5000 kilometrów sześciennych, opada na marginalne i słabo zaludnione terytoria północne.
Środowisko klimatyczne jest ważnym czynnikiem determinującym rozwój różnych gatunków flory i fauny oraz jej żyzność. Charakterystyczną cechą Rosji jest to, że większość jej terytorium ma znacznie chłodniejszy klimat niż w innych krajach.
Wszystkie rozważane komponenty środowiska są zawarte w
BIOSFERA: powłoka Ziemi, w tym część atmosfery, hydrosfera i górna część litosfery, które są połączone złożonymi cyklami biochemicznymi migracji materii i energii, powłoka geologiczna Ziemi, zamieszkana przez organizmy żywe. Górna granica życia biosfery jest ograniczona przez intensywne stężenie promieni ultrafioletowych; niższa - wysoka temperatura wnętrza ziemi (powyżej 100`C). Jego skrajne granice osiągają tylko organizmy niższe - bakterie.
Adaptację (adaptację) rośliny do określonych warunków środowiskowych zapewniają mechanizmy fizjologiczne (adaptacja fizjologiczna), aw populacji organizmów (gatunków) - mechanizmy zmienności genetycznej, dziedziczności i selekcji (adaptacja genetyczna). Czynniki środowiskowe mogą zmieniać się regularnie i losowo. Regularnie zmieniające się warunki środowiskowe (zmiana pór roku) rozwijają w roślinach adaptację genetyczną do tych warunków.
W naturalnych warunkach wzrostu lub uprawy gatunku, w trakcie swojego wzrostu i rozwoju, często doświadczają one wpływu niekorzystnych czynników środowiskowych, do których należą wahania temperatury, susza, nadmierna wilgoć, zasolenie gleby itp. Każda roślina ma umiejętność adaptacji do zmieniających się warunków środowiskowych w granicach wyznaczonych przez jego genotyp. Im wyższa zdolność rośliny do zmiany metabolizmu zgodnie ze środowiskiem, tym większa szybkość reakcji tej rośliny i lepsza zdolność adaptacji. Ta właściwość wyróżnia odporne odmiany upraw rolniczych. Z reguły niewielkie i krótkotrwałe zmiany czynników środowiskowych nie prowadzą do istotnych zaburzeń funkcji fizjologicznych roślin, co wynika z ich zdolności do utrzymywania względnie stabilnego stanu w zmieniających się warunkach środowiskowych, czyli utrzymania homeostazy. Jednak ostre i długotrwałe uderzenia prowadzą do zakłócenia wielu funkcji rośliny, a często do jej śmierci.
Pod wpływem niesprzyjających warunków obniżenie procesów i funkcji fizjologicznych może osiągnąć poziomy krytyczne, które nie zapewniają realizacji genetycznego programu ontogenezy, zaburzony zostaje metabolizm energetyczny, układy regulacyjne, metabolizm białek i inne funkcje życiowe organizmu roślinnego. Kiedy roślina jest narażona na niekorzystne czynniki (stresory), powstaje w niej stan stresu, odchylenie od normy - stres. Stres to ogólna nieswoista reakcja adaptacyjna organizmu na działanie jakichkolwiek niekorzystnych czynników. Istnieją trzy główne grupy czynników wywołujących stres u roślin: fizyczne - niedostateczna lub nadmierna wilgotność, światło, temperatura, promieniowanie radioaktywne, stres mechaniczny; chemiczne - sole, gazy, ksenobiotyki (herbicydy, insektycydy, fungicydy, odpady przemysłowe itp.); biologiczne - uszkodzenia przez patogeny lub szkodniki, konkurencja z innymi roślinami, wpływ zwierząt, kwitnienie, dojrzewanie owoców.
Reakcje na niekorzystne czynniki środowiskowe tylko w określonych warunkach są szkodliwe dla organizmów żywych iw większości przypadków mają wartość adaptacyjną. Dlatego te odpowiedzi Selye nazwał „ogólnym zespołem adaptacyjnym”. W późniejszych pracach używał terminów „stres” i „ogólny zespół adaptacyjny” jako synonimy.
Dostosowanie- jest to genetycznie zdeterminowany proces tworzenia systemów ochronnych zapewniających wzrost stabilności i przepływ ontogenezy w niesprzyjających jej warunkach.
Adaptacja jest jednym z najważniejszych mechanizmów zwiększających stabilność systemu biologicznego, w tym organizmu roślinnego, w zmienionych warunkach bytowania. Im lepiej organizm jest przystosowany do jakiegoś czynnika, tym bardziej jest odporny na jego wahania.
Określona genotypowo zdolność organizmu do zmiany metabolizmu w określonych granicach, w zależności od działania środowiska zewnętrznego, nazywana jest szybkość reakcji. Jest kontrolowany przez genotyp i jest charakterystyczny dla wszystkich żywych organizmów. Większość modyfikacji zachodzących w granicach normy reakcji ma znaczenie adaptacyjne. Odpowiadają one zmianom w siedlisku i zapewniają lepsze przetrwanie roślin w zmiennych warunkach środowiskowych. W związku z tym takie modyfikacje mają znaczenie ewolucyjne. Termin „szybkość reakcji” został wprowadzony przez V.L. Johansena (1909).
Im większa zdolność gatunku lub odmiany do modyfikacji zgodnie ze środowiskiem, tym większa szybkość reakcji i wyższa zdolność adaptacji. Ta właściwość wyróżnia odporne odmiany upraw rolniczych. Z reguły niewielkie i krótkotrwałe zmiany czynników środowiskowych nie prowadzą do znaczących naruszeń funkcji fizjologicznych roślin. Wynika to z ich zdolności do utrzymania względnej równowagi dynamicznej środowiska wewnętrznego oraz stabilności podstawowych funkcji fizjologicznych w zmieniającym się środowisku zewnętrznym. Jednocześnie ostre i długotrwałe uderzenia prowadzą do zakłócenia wielu funkcji rośliny, a często do jej śmierci.
Adaptacja obejmuje wszystkie procesy i adaptacje (anatomiczne, morfologiczne, fizjologiczne, behawioralne itp.), które zwiększają stabilność i przyczyniają się do przetrwania gatunku.
1.Adaptacje anatomiczne i morfologiczne. U niektórych przedstawicieli kserofitów długość systemu korzeniowego sięga kilkudziesięciu metrów, co pozwala roślinie korzystać z wód gruntowych i nie odczuwać braku wilgoci w warunkach glebowych i suszy atmosferycznej. U innych kserofitów obecność grubego naskórka, pokwitanie liści i przekształcenie liści w kolce ograniczają utratę wody, co jest bardzo ważne w warunkach braku wilgoci.
Płonące włosy i kolce chronią rośliny przed zjedzeniem przez zwierzęta.
Drzewa w tundrze lub na wysokich górach wyglądają jak przysadziste, płożące się krzewy, zimą pokryte są śniegiem, który chroni je przed silnymi mrozami.
W regionach górskich o dużych dobowych wahaniach temperatury rośliny często mają postać spłaszczonych poduszek z gęsto rozmieszczonymi licznymi łodygami. Pozwala to na utrzymanie wilgoci wewnątrz poduszek i względnie jednolitej temperatury przez cały dzień.
W roślinach bagiennych i wodnych tworzy się specjalny miąższ powietrznonośny (aerenchyma), który jest rezerwuarem powietrza i ułatwia oddychanie zanurzonym w wodzie częściom roślin.
2. Adaptacje fizjologiczne i biochemiczne. W przypadku sukulentów adaptacją do uprawy w warunkach pustynnych i półpustynnych jest asymilacja CO 2 podczas fotosyntezy na szlaku CAM. Te rośliny mają zamknięte aparaty szparkowe w ciągu dnia. W ten sposób roślina chroni wewnętrzne rezerwy wody przed parowaniem. Na pustyniach głównym czynnikiem ograniczającym wzrost roślin jest woda. Szparki otwierają się w nocy i w tym czasie CO 2 dostaje się do tkanek fotosyntetyzujących. Późniejsze zaangażowanie CO2 w cykl fotosyntezy następuje w ciągu dnia już przy zamkniętych aparatach szparkowych.
Adaptacje fizjologiczne i biochemiczne obejmują zdolność aparatów szparkowych do otwierania i zamykania w zależności od warunków zewnętrznych. Synteza w komórkach kwasu abscysynowego, proliny, białek ochronnych, fitoaleksyn, fitoncydów, wzrost aktywności enzymów przeciwdziałających rozpadowi oksydacyjnemu substancji organicznych, akumulacja cukrów w komórkach oraz szereg innych zmian w metabolizmie przyczynia się do wzrost odporności roślin na niekorzystne warunki środowiskowe.
Ta sama reakcja biochemiczna może być przeprowadzona przez kilka form molekularnych tego samego enzymu (izoenzymów), przy czym każda izoforma wykazuje aktywność katalityczną w stosunkowo wąskim zakresie jakiegoś parametru środowiskowego, takiego jak temperatura. Obecność szeregu izoenzymów pozwala roślinie na przeprowadzenie reakcji w znacznie szerszym zakresie temperatur w porównaniu z każdym pojedynczym izoenzymem. Dzięki temu roślina może z powodzeniem wykonywać funkcje życiowe w zmieniających się warunkach temperaturowych.
3. Adaptacje behawioralne, czyli unikanie niekorzystnego czynnika. Przykładem są efemerydy i efemerydy (mak, gwiaździsty, krokusy, tulipany, przebiśniegi). Cały cykl rozwoju przechodzą wiosną przez 1,5-2 miesiące, jeszcze przed nadejściem upałów i suszy. W ten sposób odchodzą lub unikają wpadania pod wpływ stresora. W podobny sposób wcześnie dojrzewające odmiany roślin uprawnych tworzą plon przed wystąpieniem niekorzystnych zjawisk sezonowych: sierpniowych mgły, deszczy, przymrozków. Dlatego selekcja wielu roślin uprawnych ma na celu stworzenie odmian wcześnie dojrzałych. Rośliny wieloletnie zimują jako kłącza i cebulki w glebie pod śniegiem, co chroni je przed zamarzaniem.
Adaptacja roślin do niekorzystnych czynników odbywa się jednocześnie na wielu poziomach regulacji – od pojedynczej komórki do fitocenozy. Im wyższy poziom organizacji (komórka, organizm, populacja), tym większa liczba mechanizmów jednocześnie zaangażowanych w adaptację roślin do stresu.
Regulacja procesów metabolicznych i adaptacyjnych wewnątrz komórki odbywa się za pomocą systemów: metabolicznego (enzymatycznego); genetyczny; membrana. Systemy te są ze sobą ściśle powiązane. Tak więc właściwości błon zależą od aktywności genów, a zróżnicowana aktywność samych genów jest pod kontrolą błon. Synteza enzymów i ich aktywność kontrolowana jest na poziomie genetycznym, jednocześnie enzymy regulują metabolizm kwasów nukleinowych w komórce.
Na poziom organizmu do komórkowych mechanizmów adaptacji dodawane są nowe, odzwierciedlające interakcję narządów. W niesprzyjających warunkach rośliny tworzą i zachowują taką liczbę elementów owocowych, które są dostarczane w wystarczających ilościach z niezbędnymi substancjami do wytworzenia pełnoprawnych nasion. Na przykład w kwiatostanach uprawnych zbóż i koronach drzew owocowych w niesprzyjających warunkach może odpaść ponad połowa złożonych jajników. Takie zmiany opierają się na konkurencyjnych relacjach między organami o fizjologicznie aktywne i składniki odżywcze.
W warunkach stresowych procesy starzenia i opadania dolnych liści ulegają gwałtownemu przyspieszeniu. Jednocześnie substancje niezbędne dla roślin przenoszą się z nich do młodych organów, odpowiadając na strategię przetrwania organizmu. Dzięki recyklingowi składników odżywczych z dolnych liści młodsze, górne liście, pozostają żywotne.
Istnieją mechanizmy regeneracji utraconych narządów. Na przykład powierzchnia rany pokryta jest wtórną tkanką powłokową (perydermą rany), rana na pniu lub gałęzi goi się napływami (modzele). Wraz z utratą pędu wierzchołkowego w roślinach budzą się uśpione pąki i intensywnie rozwijają się pędy boczne. Wiosenne odtworzenie liści zamiast opadłych jesienią jest również przykładem naturalnej regeneracji narządów. Regeneracja jako urządzenie biologiczne, które zapewnia rozmnażanie wegetatywne roślin przez segmenty korzeni, kłącza, plechy, sadzonki łodyg i liści, izolowane komórki, pojedyncze protoplasty, ma ogromne znaczenie praktyczne w uprawie roślin, sadownictwie, leśnictwie, ogrodnictwie ozdobnym itp.
Układ hormonalny bierze również udział w procesach ochrony i adaptacji na poziomie roślin. Na przykład pod wpływem niesprzyjających warunków w roślinie gwałtownie wzrasta zawartość inhibitorów wzrostu: etylenu i kwasu abscysynowego. Ograniczają przemianę materii, hamują procesy wzrostu, przyspieszają starzenie się, upadek narządów i przejście rośliny w stan uśpienia. Charakterystyczną reakcją roślin jest zahamowanie czynności funkcjonalnej pod wpływem stresu pod wpływem inhibitorów wzrostu. Jednocześnie w tkankach zmniejsza się zawartość stymulatorów wzrostu: cytokininy, auksyny i giberelin.
Na poziom populacji dodaje się selekcję, co prowadzi do pojawienia się bardziej przystosowanych organizmów. O możliwości selekcji decyduje występowanie wewnątrzpopulacyjnej zmienności odporności roślin na różne czynniki środowiskowe. Przykładem wewnątrzpopulacyjnej zmienności odporności może być nieprzyjazne pojawianie się siewek na zasolonej glebie oraz zwiększenie zmienności czasu kiełkowania wraz ze wzrostem działania stresora.
Gatunek we współczesnym ujęciu składa się z dużej liczby biotypów - mniejszych jednostek ekologicznych, identycznych genetycznie, ale wykazujących różną odporność na czynniki środowiskowe. W różnych warunkach nie wszystkie biotypy są jednakowo ważne, aw wyniku konkurencji pozostają tylko te, które najlepiej spełniają dane warunki. Oznacza to, że odporność populacji (odmiany) na określony czynnik zależy od odporności organizmów tworzących populację. Odmiany odporne mają w swoim składzie zestaw biotypów, które zapewniają dobrą wydajność nawet w niesprzyjających warunkach.
Jednocześnie w procesie wieloletniej uprawy skład i proporcja biotypów w populacji zmienia się w odmianach, co wpływa na produktywność i jakość odmiany, często nie na lepsze.
Tak więc adaptacja obejmuje wszystkie procesy i adaptacje, które zwiększają odporność roślin na niekorzystne warunki środowiskowe (anatomiczne, morfologiczne, fizjologiczne, biochemiczne, behawioralne, populacyjne itp.)
Ale aby wybrać najskuteczniejszy sposób adaptacji, najważniejszy jest czas, w którym organizm musi przystosować się do nowych warunków.
Przy nagłym działaniu czynnika ekstremalnego reakcja nie może być opóźniona, musi nastąpić natychmiast, aby wykluczyć nieodwracalne uszkodzenie rośliny. Przy długotrwałych skutkach małej siły, adaptacyjne przegrupowania następują stopniowo, podczas gdy wybór możliwych strategii rośnie.
W związku z tym istnieją trzy główne strategie adaptacyjne: ewolucyjny, ontogenetyczny oraz pilne. Zadaniem strategii jest efektywne wykorzystanie dostępnych zasobów do osiągnięcia głównego celu – przetrwania organizmu w stresie. Strategia adaptacyjna ma na celu utrzymanie integralności strukturalnej ważnych makrocząsteczek i czynności funkcjonalnej struktur komórkowych, utrzymanie systemów regulacji aktywności życiowej oraz dostarczanie roślinom energii.
Adaptacje ewolucyjne lub filogenetyczne(filogeneza - rozwój gatunku biologicznego w czasie) - są to adaptacje, które powstają w procesie ewolucyjnym na podstawie mutacji genetycznych, selekcji i są dziedziczone. Są najbardziej niezawodne w przetrwaniu roślin.
Każdy gatunek roślin w procesie ewolucji rozwinął pewne potrzeby dotyczące warunków istnienia i przystosowania do zajmowanej niszy ekologicznej, stabilnej adaptacji organizmu do środowiska. Tolerancja na wilgoć i cień, ciepło, zimno i inne cechy ekologiczne poszczególnych gatunków roślin ukształtowały się w wyniku długotrwałego działania odpowiednich warunków. Tak więc rośliny ciepłolubne i krótkodniowe są charakterystyczne dla południowych szerokości geograficznych, mniej wymagające ciepła i rośliny o długim dniu są charakterystyczne dla północnych szerokości geograficznych. Dobrze znane są liczne ewolucyjne adaptacje kserofitów do suszy: ekonomiczne wykorzystanie wody, głęboki system korzeniowy, zrzucanie liści i przejście w stan uśpienia oraz inne adaptacje.
Pod tym względem odmiany roślin rolniczych wykazują odporność właśnie na te czynniki środowiskowe, przeciwko którym prowadzona jest hodowla i selekcja form produkcyjnych. Jeśli selekcja odbywa się w kilku kolejnych pokoleniach na tle stałego wpływu jakiegoś niekorzystnego czynnika, to odporność odmiany na nią może zostać znacznie zwiększona. Naturalne jest, że odmiany wyhodowane przez Instytut Badawczy Rolnictwa Południowego Wschodu (Saratow) są bardziej odporne na suszę niż odmiany wyhodowane w ośrodkach hodowlanych regionu moskiewskiego. W ten sam sposób w strefach ekologicznych o niesprzyjających warunkach glebowo-klimatycznych wykształciły się odporne lokalne odmiany roślin, a gatunki endemiczne są odporne na stresor wyrażający się w ich siedlisku.
Charakterystyka odporności odmian pszenicy jarej z kolekcji Wszechrosyjskiego Instytutu Przemysłu Roślinnego (Semenov i in., 2005)
| Różnorodność | Początek | Zrównoważony rozwój |
| Enita | region Moskwy | Średnio odporny na suszę |
| Saratowska 29 | obwód saratowski | odporny na suszę |
| Kometa | Region Swierdłowska. | odporny na suszę |
| Karazin | Brazylia | kwasoodporny |
| Preludium | Brazylia | kwasoodporny |
| Kolonias | Brazylia | kwasoodporny |
| Thrintani | Brazylia | kwasoodporny |
| PPG-56 | Kazachstan | tolerancja na sól |
| Osz | Kirgistan | tolerancja na sól |
| Surkhak 5688 | Tadżykistan | tolerancja na sól |
| Messel | Norwegia | Tolerancja na sól |
W środowisku naturalnym warunki środowiskowe zwykle zmieniają się bardzo szybko, a czas, w którym czynnik stresowy osiąga szkodliwy poziom, nie wystarcza do powstania adaptacji ewolucyjnych. W takich przypadkach rośliny wykorzystują nie trwałe, ale wywołane stresorami mechanizmy obronne, których powstanie jest zdeterminowane (zdeterminowane) genetycznie.
Adaptacje ontogenetyczne (fenotypowe) nie są związane z mutacjami genetycznymi i nie są dziedziczone. Powstawanie takich adaptacji wymaga stosunkowo długiego czasu, dlatego nazywane są adaptacjami długotrwałymi. Jednym z takich mechanizmów jest zdolność wielu roślin do tworzenia wodooszczędnej ścieżki fotosyntezy typu CAM w warunkach deficytu wody spowodowanego suszą, zasoleniem, niskimi temperaturami i innymi stresorami.
Adaptacja ta związana jest z indukcją ekspresji genu karboksylazy fosfoenolopirogronianowej, który w normalnych warunkach jest nieaktywny, oraz genów innych enzymów szlaku wychwytywania CO2 CAM, z biosyntezą osmolitów (proliny), z aktywacją przeciwutleniacza systemy i zmiany w dziennym rytmie ruchów szparkowych. Wszystko to prowadzi do bardzo ekonomicznego zużycia wody.
W uprawach polowych, na przykład w kukurydzy, aerenchyma nie występuje w normalnych warunkach wzrostu. Jednak w warunkach powodzi i braku tlenu w tkankach w korzeniach, niektóre komórki pierwotnej kory korzenia i łodygi obumierają (apoptoza lub zaprogramowana śmierć komórki). Na ich miejscu powstają wnęki, przez które tlen jest transportowany z nadziemnych części rośliny do systemu korzeniowego. Sygnałem do śmierci komórki jest synteza etylenu.
Pilna adaptacja występuje z szybkimi i intensywnymi zmianami warunków życia. Opiera się na tworzeniu i funkcjonowaniu systemów ochrony przed wstrząsami. Do systemów obrony przed wstrząsami zalicza się na przykład białkowy system szoku cieplnego, który powstaje w odpowiedzi na gwałtowny wzrost temperatury. Mechanizmy te zapewniają krótkoterminowe warunki do przetrwania pod działaniem czynnika uszkadzającego, a tym samym stwarzają warunki do powstania bardziej niezawodnych, długoterminowych wyspecjalizowanych mechanizmów adaptacyjnych. Przykładem wyspecjalizowanych mechanizmów adaptacyjnych jest tworzenie się białek przeciw zamarzaniu w niskich temperaturach lub synteza cukrów podczas przezimowania upraw ozimych. Jednocześnie, jeśli szkodliwy wpływ czynnika przekracza ochronne i naprawcze możliwości organizmu, nieuchronnie następuje śmierć. W tym przypadku organizm umiera na etapie nagłego lub na etapie adaptacji specjalistycznej, w zależności od intensywności i czasu trwania czynnika ekstremalnego.
Wyróżnić konkretny oraz niespecyficzne (ogólne) reakcje roślin na stresory.
Reakcje niespecyficzne nie zależą od charakteru działającego czynnika. Są takie same pod wpływem wysokich i niskich temperatur, braku lub nadmiaru wilgoci, wysokiego stężenia soli w glebie czy szkodliwych gazów w powietrzu. We wszystkich przypadkach zwiększa się przepuszczalność błon w komórkach roślinnych, zaburza się oddychanie, nasila się hydrolityczny rozkład substancji, wzrasta synteza etylenu i kwasu abscysynowego, zahamowany jest podział i wydłużanie komórek.
W tabeli przedstawiono zespół niespecyficznych zmian zachodzących w roślinach pod wpływem różnych czynników środowiskowych.
Zmiany parametrów fizjologicznych roślin pod wpływem warunków stresowych (wg G.V., Udovenko, 1995)
| Opcje | Charakter zmiany parametrów w warunkach | |||
| susze | zasolenie | wysoka temperatura | niska temperatura | |
| Stężenie jonów w tkankach | rozwój | rozwój | rozwój | rozwój |
| Aktywność wody w komórce | Spadanie w dół | Spadanie w dół | Spadanie w dół | Spadanie w dół |
| Potencjał osmotyczny komórki | rozwój | rozwój | rozwój | rozwój |
| Pojemność wodna | rozwój | rozwój | rozwój | — |
| Niedobór wody | rozwój | rozwój | rozwój | — |
| Przepuszczalność protoplazmy | rozwój | rozwój | rozwój | — |
| Szybkość transpiracji | Spadanie w dół | Spadanie w dół | rozwój | Spadanie w dół |
| Wydajność transpiracji | Spadanie w dół | Spadanie w dół | Spadanie w dół | Spadanie w dół |
| Efektywność energetyczna oddychania | Spadanie w dół | Spadanie w dół | Spadanie w dół | — |
| Intensywność oddychania | rozwój | rozwój | rozwój | — |
| Fotofosforylacja | Zmniejsza | Zmniejsza | — | Zmniejsza |
| Stabilizacja jądrowego DNA | rozwój | rozwój | rozwój | rozwój |
| Funkcjonalna aktywność DNA | Zmniejsza | Zmniejsza | Zmniejsza | Zmniejsza |
| Koncentracja proliny | rozwój | rozwój | rozwój | — |
| Zawartość białek rozpuszczalnych w wodzie | rozwój | rozwój | rozwój | rozwój |
| Reakcje syntetyczne | Zduszony | Zduszony | Zduszony | Zduszony |
| Pobieranie jonów przez korzenie | Zduszony | Zduszony | Zduszony | Zduszony |
| Transport substancji | W depresji | W depresji | W depresji | W depresji |
| Koncentracja pigmentu | Spadanie w dół | Spadanie w dół | Spadanie w dół | Spadanie w dół |
| podział komórek | zwalnia | zwalnia | — | — |
| Rozciąganie komórek | Zduszony | Zduszony | — | — |
| Liczba elementów owocowych | Zredukowany | Zredukowany | Zredukowany | Zredukowany |
| Starzenie się narządów | Przyśpieszony | Przyśpieszony | Przyśpieszony | — |
| żniwa biologiczne | Obniżony | Obniżony | Obniżony | Obniżony |
Na podstawie danych zawartych w tabeli można zauważyć, że odporności roślin na kilka czynników towarzyszą jednokierunkowe zmiany fizjologiczne. Daje to powody sądzić, że wzrostowi odporności roślin na jeden czynnik może towarzyszyć wzrost odporności na inny. Zostało to potwierdzone eksperymentami.
Eksperymenty w Instytucie Fizjologii Roślin Rosyjskiej Akademii Nauk (Vl. V. Kuznetsov i inni) wykazały, że krótkotrwałej obróbce cieplnej roślin bawełny towarzyszy wzrost ich odporności na późniejsze zasolenie. A przystosowanie roślin do zasolenia prowadzi do wzrostu ich odporności na wysokie temperatury. Szok cieplny zwiększa zdolność roślin do przystosowania się do następującej po niej suszy i odwrotnie, w procesie suszy wzrasta odporność organizmu na wysoką temperaturę. Krótkotrwała ekspozycja na wysokie temperatury zwiększa odporność na metale ciężkie i promieniowanie UV-B. Poprzednia susza sprzyja przetrwaniu roślin w warunkach zasolenia lub chłodu.
Proces zwiększania odporności organizmu na dany czynnik środowiskowy w wyniku adaptacji do czynnika o innym charakterze nazywa się krzyżowa adaptacja.
W badaniu ogólnych (niespecyficznych) mechanizmów odporności bardzo interesująca jest reakcja roślin na czynniki powodujące niedobór wody w roślinach: zasolenie, suszę, niskie i wysokie temperatury oraz kilka innych. Na poziomie całego organizmu wszystkie rośliny w ten sam sposób reagują na niedobór wody. Charakteryzuje się zahamowaniem wzrostu pędów, zwiększonym wzrostem systemu korzeniowego, syntezą kwasu abscysynowego oraz spadkiem przewodnictwa szparkowego. Po pewnym czasie dolne liście szybko się starzeją i obserwuje się ich obumieranie. Wszystkie te reakcje mają na celu zmniejszenie zużycia wody poprzez zmniejszenie powierzchni parowania, a także zwiększenie aktywności absorpcyjnej korzenia.
Reakcje specyficzne są reakcjami na działanie jednego czynnika stresowego. Tak więc fitoaleksyny (substancje o właściwościach antybiotycznych) są syntetyzowane w roślinach w odpowiedzi na kontakt z patogenami (patogenami).
Specyfika lub niespecyficzność odpowiedzi implikuje z jednej strony stosunek rośliny do różnych stresorów, az drugiej charakterystyczne reakcje roślin różnych gatunków i odmian na ten sam stresor.
Przejawy specyficznych i niespecyficznych odpowiedzi roślin zależą od siły stresu i tempa jego rozwoju. Specyficzne reakcje pojawiają się częściej, jeśli stres rozwija się powoli, a organizm ma czas na odbudowę i przystosowanie się do niego. Reakcje niespecyficzne występują zwykle przy krótszym i silniejszym działaniu stresora. Funkcjonowanie niespecyficznych (ogólnych) mechanizmów odpornościowych pozwala roślinie uniknąć dużych nakładów energetycznych na tworzenie wyspecjalizowanych (specyficznych) mechanizmów adaptacyjnych w odpowiedzi na wszelkie odchylenia od normy w ich warunkach życia.
Odporność roślin na stres zależy od fazy ontogenezy. Najbardziej stabilne rośliny i organy roślinne w stanie uśpienia: w postaci nasion, cebulek; zdrewniałe byliny – po opadnięciu liści w stanie głębokiego spoczynku. Rośliny są najbardziej wrażliwe w młodym wieku, ponieważ procesy wzrostu ulegają zniszczeniu w pierwszej kolejności w warunkach stresowych. Drugi okres krytyczny to okres tworzenia gamet i zapłodnienia. Efekt stresu w tym okresie prowadzi do zmniejszenia funkcji rozrodczej roślin i spadku plonu.
Jeśli warunki stresowe powtarzają się i mają niską intensywność, przyczyniają się do twardnienia roślin. Na tej podstawie powstają metody zwiększania odporności na niskie temperatury, ciepło, zasolenie oraz zwiększoną zawartość szkodliwych gazów w powietrzu.
Niezawodność organizmu roślinnego zależy od jego zdolności do zapobiegania lub eliminowania awarii na różnych poziomach organizacji biologicznej: molekularnej, subkomórkowej, komórkowej, tkankowej, narządowej, organizmu i populacji.
Aby zapobiec zakłóceniom w życiu roślin pod wpływem niekorzystnych czynników, zasady nadmierność, niejednorodność składników funkcjonalnie równoważnych, systemy do naprawy utraconych konstrukcji,.
Redundancja struktur i funkcjonalności to jeden z głównych sposobów zapewnienia niezawodności systemów. Nadmiarowość i nadmiarowość ma wiele przejawów. Na poziomie subkomórkowym rezerwacja i powielanie materiału genetycznego przyczynia się do wzrostu niezawodności organizmu roślinnego. Zapewnia to na przykład podwójna helisa DNA poprzez zwiększenie ploidii. Niezawodność funkcjonowania organizmu roślinnego w zmieniających się warunkach jest również wspierana przez obecność różnych cząsteczek informacyjnego RNA oraz powstawanie heterogenicznych polipeptydów. Należą do nich izoenzymy, które katalizują tę samą reakcję, ale różnią się właściwościami fizykochemicznymi i stabilnością struktury molekularnej w zmieniających się warunkach środowiskowych.
Na poziomie komórkowym przykładem nadmiarowości jest nadmiar organelli komórkowych. W ten sposób ustalono, że część dostępnych chloroplastów wystarcza do dostarczenia roślinie produktów fotosyntezy. Pozostałe chloroplasty niejako pozostają w rezerwie. To samo dotyczy całkowitej zawartości chlorofilu. Redundancja objawia się również dużym nagromadzeniem prekursorów biosyntezy wielu związków.
Na poziomie organizmu zasada redundancji wyraża się w tworzeniu i układaniu w różnym czasie większej ilości pędów, kwiatów, kłosków niż jest to wymagane do zmiany pokoleń, w ogromnej ilości pyłku, zalążków, nasion.
Na poziomie populacji zasada redundancji przejawia się w dużej liczbie osobników różniących się odpornością na określony czynnik stresowy.
Systemy naprawcze działają również na różnych poziomach - molekularnym, komórkowym, organizmalnym, populacyjnym i biocenotycznym. Procesy naprawcze przebiegają wraz z wydatkowaniem energii i substancji plastycznych, dlatego naprawa jest możliwa tylko wtedy, gdy utrzymane jest wystarczające tempo przemiany materii. Jeśli przemiana materii ustaje, ustaje również regeneracja. W ekstremalnych warunkach środowiska zewnętrznego zachowanie oddychania jest szczególnie ważne, ponieważ oddychanie dostarcza energii do procesów naprawczych.
Zdolność regeneracyjna komórek organizmów zaadaptowanych jest determinowana przez odporność ich białek na denaturację, czyli stabilność wiązań, które determinują drugorzędową, trzeciorzędową i czwartorzędową strukturę białka. Na przykład odporność dojrzałych nasion na wysokie temperatury jest zwykle związana z tym, że po odwodnieniu ich białka stają się odporne na denaturację.
Głównym źródłem materiału energetycznego jako substratu do oddychania jest fotosynteza, dlatego zaopatrzenie komórki w energię i związane z tym procesy naprawcze zależą od stabilności i zdolności aparatu fotosyntetycznego do regeneracji po uszkodzeniu. Aby utrzymać fotosyntezę w ekstremalnych warunkach w roślinach, aktywowana jest synteza składników błony tylakoidowej, hamowane jest utlenianie lipidów i przywracana jest ultrastruktura plastydu.
Na poziomie organizmu przykładem regeneracji jest rozwój pędów zastępczych, przebudzenie uśpionych pąków w przypadku uszkodzenia punktów wzrostu.
Jeśli znajdziesz błąd, zaznacz fragment tekstu i kliknij Ctrl+Enter.
(według N. Greena i in., 1993)
| Dostosowanie | Przykłady | ||
| Zmniejszenie utraty wody | |||
| Liście zamienione w igły lub kolce Zanurzone szparki Liście zwinięte w cylinder Gruba woskowa łuska Gruba łodyga o wysokim stosunku objętości do powierzchni Owłosione liście Zrzucanie liści podczas suszy Szparki otwarte na noc i zamknięte w ciągu dnia Kwasy Wydłużona pętla Henle w nerki Tkanki tolerują ciepło ze względu na zmniejszoną potliwość lub transpirację Zwierzęta chowają się w norach Otwory oddechowe są zakryte zaworami | Cactaceae, Euphorbiaceae (euphorbiaceae), drzewa iglaste Pinus, Ammophila Ammophila Liście większości kserofitów, owady Cactaceae, Euphorbiaceae („sukulenty”) Wiele roślin alpejskich Fouguieria splendens Crassulaceae (grubożerne, pospolite, niektóre gatunki ptaków pustynnych) C-4. ssaki, np. wielbłąd, szczur pustynny Wiele roślin pustynnych, wielbłąd Wiele małych ssaków pustynnych, np. szczur pustynny Wiele owadów | ||
| Zwiększ wchłanianie wody | |||
| Rozległy płytki system korzeniowy i głęboko penetrujące korzenie Długie korzenie Kopanie przejść do wody | Niektóre Cactaceae, takie jak Opuntia i Euphorbiaceae Wiele roślin alpejskich, takich jak szarotka alpejska (Leontopodium alpinum) Termity | ||
| magazynowanie wody | |||
| W komórkach śluzowych i ścianach komórkowych W wyspecjalizowanym pęcherzu W postaci tłuszczu (woda jest produktem utleniania) | Cactaceae i Euphorbiaceae Żaba pustynna Szczur pustynny | ||
| Fizjologiczna odporność na utratę wody | |||
| Widoczne odwodnienie pozostaje możliwe Utrata znacznej części masy ciała i szybki powrót do zdrowia dzięki dostępnej wodzie | Niektóre paprocie epifityczne i widłaki, wiele mszaków i porostów, turzyca Carex physoides Lumbricus terrestris (traci do 70% masy), wielbłąd (traci do 30%) | ||
Koniec tabeli 4,9
Połączone działanie temperatury
I wilgotność
Uwzględnienie poszczególnych czynników środowiskowych nie jest ostatecznym celem badań ekologicznych, ale sposobem podejścia do złożonych problemów środowiskowych, dać porównawczą ocenę znaczenia różnych czynników współdziałających w rzeczywistych ekosystemach.
Temperatura i wilgotność są wiodącymi czynnikami klimatycznymi i są ze sobą ściśle powiązane (ryc. 4.19).
Ryż. 4.19. Wpływ temperatury na wilgotność względną
powietrze (według B. Nebla, 1993)
Przy stałej ilości wody w powietrzu wilgotność względna wzrasta wraz ze spadkiem temperatury. Jeśli powietrze zostanie schłodzone poniżej punktu nasycenia wodą (100%), zachodzi kondensacja i następuje opady. Po podgrzaniu jego wilgotność względna spada. Połączenie temperatury i wilgotności często odgrywa decydującą rolę w rozmieszczeniu roślinności i zwierząt. Interakcja temperatury i wilgotności zależy nie tylko od względnej, ale także od ich wartości bezwzględnej. Na przykład temperatura ma bardziej wyraźny wpływ na organizmy w warunkach wilgotności zbliżonej do krytycznej, to znaczy, gdy wilgotność jest bardzo wysoka lub bardzo niska. Wilgotność odgrywa również bardziej krytyczną rolę w temperaturach zbliżonych do ekstremalnych. Dlatego te same gatunki organizmów na różnych obszarach geograficznych preferują różne siedliska. Tak, do zasada wstępna, ustanowiony przez V. V. Alekhina (1951) dla roślinności, na północnych zboczach rosną szeroko rozpowszechnione gatunki na południu, a na północy występują tylko na południowych (ryc. 4.20).
Ryż. 4.20. Schemat reguły antycypacji (według V.V. Alechina, 1951):
1 - gatunki północne żyjące na wyżynie, od południa przechodzące na stoki ekspozycji północnej i w belki; 2 - gatunek południowy, występujący na północy na najcieplejszych stokach o ekspozycji południowej
Zasady ujawnione dla zwierząt zmiana siedliska(G. Ya. Bei-Bienko, 1961) i zasada zmiany poziomów(M. S. Gilyarov, 1970), gdzie gatunki mezofilne znajdują się w centrum zasięgu, na północy wybierają miejsca bardziej suche, a na południu - miejsca bardziej wilgotne lub przechodzą z trybu życia naziemnego do podziemnego, jak wiele fitofagów. Im słabszy wpływ klimatu przejawia się w konkretnych siedliskach, które wybiera gatunek, tym większa jest jego zdolność do życia w różnych warunkach klimatycznych. Gatunek wybiera kombinację czynników najbardziej odpowiednią dla swojej wartościowości ekologicznej poprzez zmianę swojego siedliska, a tym samym pokonuje granice klimatyczne.
Zależność między temperaturą a wilgotnością dobrze odzwierciedlają wykresy klimatyczne opracowane według metoda Walthera-Gossena, na którym, w określonej skali, porównuje się roczny przebieg temperatury powietrza z przebiegiem opadów (ryc. 4.21).

Ryż. 4.21. Wykres klimatyczny według Waltera-Gossena dla Odessy
(według G. Waltera, 1968):
a - wysokość nad poziomem morza; b - liczba lat obserwacji temperatury (pierwsza cyfra) i opadów (druga cyfra); c - średnia roczna temperatura; d - średnie roczne opady w mm d - średnie minimum dobowe najzimniejszego miesiąca; e jest absolutnym minimum; w - średnie dzienne maksimum najcieplejszego miesiąca; h - absolutne maksimum; oraz - krzywa średnich miesięcznych temperatur; k - krzywa średnich opadów miesięcznych (stosunek 10°=20 mm); l - to samo (stosunek 10°=30 mm); m - okres suchy; n - okres półpustynny; o - pora deszczowa; n - miesiące ze średnią dobową temperaturą minimalną poniżej 0°C; p - miesiące z absolutną minimalną temperaturą poniżej 0°C, c - okres bez mrozu. Na odciętej - miesiące
Wykresy klimatyczne można budować dla poszczególnych lat, a układając je sekwencyjnie i nieprzerwanie jeden po drugim, można uzyskać klimatogram. Ekstremalnie suche lub ekstremalnie zimne lata można łatwo prześledzić na klimatogramach, co jest bardzo przydatne przy określaniu przydatności kombinacji temperatury i wilgotności na obszarach, na których przewiduje się wprowadzenie roślin lub zwierząt łownych.
Atmosfera
Jak wspomniano wcześniej, nasza planeta Ziemia różni się od innych planet obecnością powłoki powietrznej, atmosfery, powietrza atmosferycznego. Powietrze atmosferyczne to mieszanina różnych gazów. Zawiera 78,08% azotu, 20,9% tlenu, 0,93% argonu, 0,03% dwutlenku węgla, inne gazy (hel, metan, neon, ksenon, radon itp.) około 0,01%.
Znaczenie powietrza atmosferycznego dla organizmów żywych jest ogromne i zróżnicowane. Jest źródłem tlenu do oddychania i dwutlenku węgla do fotosyntezy. Chroni organizmy żywe przed szkodliwym promieniowaniem kosmicznym, pomaga zachować ciepło na Ziemi.
Atmosfera jest ważną częścią ekosfery, z którą jest połączona cyklami biogeochemicznymi, w tym składnikami gazowymi. Są to obiegi węgla, azotu, tlenu i wody. Duże znaczenie mają również fizyczne właściwości atmosfery. Powietrze stawia więc tylko niewielkie opory ruchu i nie może służyć jako podpora dla organizmów lądowych, które bezpośrednio wpłynęły na ich strukturę. W tym samym czasie niektóre grupy zwierząt zaczęły wykorzystywać lot jako środek transportu. Należy szczególnie zauważyć, że w atmosferze istnieje stały obieg mas powietrza, którego energię dostarcza Słońce (ryc. 4.22).

Ryż. 4.22. Uproszczony schemat ogólnego obiegu
masy powietrza atmosfery:
1 - ciepłe powietrze; 2 - schłodzone powietrze; 3 - strefy wysokiego ciśnienia; CE - pasaty; SD - dominujące wiatry południowo-zachodnie; GH - polarne wiatry północno-wschodnie
Skutkiem cyrkulacji jest redystrybucja pary wodnej, ponieważ atmosfera wychwytuje ją w jednym miejscu (gdzie woda paruje), przenosi i oddaje w innym miejscu (gdzie opad pada). Jeżeli gazy dostaną się do atmosfery, w tym zanieczyszczenia, takie jak dwutlenek siarki na terenach przemysłowych, to system cyrkulacji atmosferycznej dokona ich redystrybucji i wyrzucą w innych miejscach rozpuszczone w wodach opadowych (rys. 4.23).
Wiatr, oddziałując z innymi czynnikami środowiskowymi, może wpływać na rozwój roślinności, przede wszystkim drzew rosnących na terenach otwartych. Zwykle powoduje to zahamowanie wzrostu i pochylenie się na wiatr.
Wiatr odgrywa ważną rolę w rozprzestrzenianiu się zarodników, nasion itp., rozszerzając możliwości rozprzestrzeniania się organizmów nieruchomych – roślin, grzybów i niektórych bakterii. Wiatr może również wpływać na migrację latających zwierząt.

Ryż. 4.23. Cykl hydrologiczny i magazynowanie wody
(według E.A. Kriksunova i in., 1995)
Inną cechą atmosfery jest jej ciśnienie, które maleje wraz z wysokością. Ewolucja organizmów żywych na naszej planecie odbywała się przy ciśnieniu atmosferycznym 760 mm Hg na poziomie morza i jest uważana za „normalną”. Wraz ze wzrostem wysokości, na przykład, gdy ludzie wspinają się po górach, może wystąpić stan z powodu niewystarczającego nasycenia krwi tlenem. niedotlenienie lub niedotlenienie. Wynika to z faktu, że wraz ze wzrostem wysokości spada ciśnienie parcjalne tlenu, a także innych gazów zawartych w powietrzu atmosferycznym. Na wysokości 5450 m ciśnienie atmosferyczne jest o połowę mniejsze niż na poziomie morza. I chociaż powietrze zawiera tutaj taki sam procent tlenu, jego stężenie na jednostkę objętości jest o połowę mniejsze.
U roślin w tych warunkach wzrasta transpiracja, co wymagało rozwoju adaptacji do oszczędzania wody, jak na przykład u wielu roślin alpejskich.
Topografia
Topografia(relief) odnosi się do czynników orograficznych i jest ściśle związany z innymi czynnikami abiotycznymi, chociaż nie należą one do tak bezpośrednich czynników środowiskowych jak światło, ciepło, woda i gleba. Głównym czynnikiem topograficznym (orograficznym) jest wysokość. Wraz z wysokością spadają średnie temperatury, wzrasta dobowa różnica temperatur, zwiększa się ilość opadów, prędkość wiatru i natężenie promieniowania, spada ciśnienie atmosferyczne i stężenie gazów. Tym samym wzrostowi poziomu terenu na każde 100 m towarzyszy spadek temperatury powietrza o około 0,6°C.
W zależności od wielkości form, topografia lub relief dzieli się na kilka rzędów: makrorelief(góry, zagłębienia międzygórskie, niziny), mezorelief(wzgórza, wąwozy, grzbiety, zapadliska, stepowe „spodki” itp.) i mikrorelief(małe zagłębienia, nierówności, wzniesienia przy pniach itp.), wszystko to ma wpływ na rośliny i zwierzęta. W rezultacie podział na strefy pionowe stał się powszechny (rysunek 4.24).

Ryż. 4.24. Schemat przedstawiający korespondencję między kolejnymi
pionowe i poziome strefy roślinności:
1 - strefa tropikalna (strefa lasów tropikalnych); 2 - strefa umiarkowana (strefa lasów liściastych i iglastych); 3 - strefa alpejska (strefa roślinności zielnej, mchów i porostów); 4 - strefa polarna (strefa śniegu i lodu)
Pasma górskie mogą służyć jako bariery klimatyczne. Wilgotne powietrze ochładza się, gdy unosi się nad górami, powodując duże ilości opadów na nawietrznych zboczach.
Po zawietrznej stronie pasma górskiego tworzy się tzw. „cień deszczu”, powietrze jest tu bardziej suche, spada mniej opadów, tworzą się warunki pustynne, ponieważ powietrze opadające nagrzewa się i pochłania wilgoć z gleby.
Wpływa na żywe organizmy. Dla większości kręgowców górna granica życia wynosi około 6,0 km. Spadek ciśnienia wraz ze wzrostem pociąga za sobą zmniejszenie dopływu tlenu i odwodnienie zwierząt z powodu wzrostu częstości oddechów. Nieco bardziej odporne są stawonogi (skoczogonki, roztocza, pająki), które można znaleźć na lodowcach powyżej granicy roślinności. Rośliny alpejskie charakteryzują się przysadzistym wzrostem. We wszystkich wysokogórskich rejonach kuli ziemskiej przeważają niewymiarowe krzewy i krzewy pełzające (ryc. 4.25), wieloletnie trawy poduszkowe i rozetowe, darniowce i turzyce, mchy i porosty.

Ryż. 4.25. Juniper Turkestan - na zboczach grzbietu
Terekei-Alatau (według IG Serebryakov, 1955):
A - forma drzewiasta (pas łąkowo-leśny, 2900 m n.p.m.); B - elfin (pas subalpejski, 3200 m n.p.m.)
Charakterystyczną cechą morfologiczną wielu roślin alpejskich przysadzistych, takich jak krzewy i krzewinki, jest znaczna przewaga masy podziemnej nad nadziemną.
Niski wzrost roślin alpejskich wiąże się z adaptacją do niskich temperatur i kształtującym działaniem promieniowania, które bogate jest w krótkofalową część widma, hamującą procesy wzrostu. W budowie anatomicznej roślin alpejskich istnieje szereg cech, które przyczyniają się do ochrony przed nadmiernym promieniowaniem, są związane z naturą reżimu wodnego i metabolizmem na wyżynach: pogrubienie tkanek powłokowych, które zapewniają odporność na silne wiatry itp. Rośliny żyjące na skałach doświadczają zmian w kierunku kseromorfozy: zmniejszają się rozmiary komórek i zwiększa się gęstość tkanek, zwiększa się liczba aparatów szparkowych na jednostkę powierzchni liścia, zmniejszają się ich rozmiary. U gatunków żyjących w pobliżu roztopów lub innych źródeł wilgoci liście są większe, a cechy kseromorficzne mniej wyraźne.
Niskie temperatury i silne światło sprzyjają tworzeniu się dużych ilości antocyjanów, stąd głębokie, bogate tony koloru kwiatów. Charakterystyczną cechą wielu roślin alpejskich jest połączenie małych liści o niewielkim wzroście z dużymi, jaskrawo ubarwionymi kwiatami.
Cechą charakterystyczną fizjologii i biochemii roślin wysokogórskich jest wzrost intensywności procesów redoks, wzrost aktywności zaangażowanych w nie enzymów (katalazy, peroksydazy itp.) oraz niższe optimum temperaturowe ich pracy niż w roślinach nizinnych.
Oddychanie roślin alpejskich jest odporne na niekorzystne działanie, z reguły obserwuje się wzrost oddychania, a co za tym idzie, wzrost energii uwalnianej podczas rozkładu związków kompleksowych. Według współczesnych koncepcji jest to jedna z fizjologicznych podstaw adaptacji roślin do ekstremalnych warunków.
Podczas wspinaczki w góry zmienia się również sezonowy rozwój roślin. Tak więc wiosną, wspinając się po górach, można zaobserwować rozwój tego samego gatunku w następującej kolejności: w strefie niskogórskiej - średnio kwitnienie - pączkowanie, jeszcze wyżej - początek wegetacji, oraz, w końcu tylko wygląd po stopieniu śniegu. Jesienią, wspinając się w góry, obserwujemy przyspieszony początek jesiennych fenofaz: zabarwienia liści, opadania liści i obumierania części nadziemnych. U roślin następuje wyraźne skrócenie okresu wegetacji.
Wraz z wysokością nad poziomem morza duże znaczenie dla organizmów żywych ma ekspozycja i stromość zboczy.
Na półkuli północnej południowe zbocza gór otrzymują więcej światła słonecznego, natężenie światła i temperatura są tu wyższe niż na dnie doliny i na zboczach ekspozycji północnej. Na półkuli południowej sytuacja jest odwrotna. Ma to uderzający wpływ zarówno na naturalną roślinność, jak i ziemie użytkowane przez człowieka. Na przykład szerokie szczeliny między skałami nad Dunajem we wschodniej Serbii, chronione przed wiatrem i doświadczające nawilżającego działania rzeki, przyczyniły się do ochrony wielu rzadkich, reliktowych i endemicznych gatunków roślin, w tym „leszczyny niedźwiedziej” – Corylus colurna, orzech - Juglans regia, liliowy (forma dzika) - Syringa vulgaris i inne.
Strome zbocza charakteryzują się szybkim odwadnianiem i erozją gleby. Tutaj gleby są zwykle cienkie i suche, z roślinnością kseromorficzną. Przy nachyleniu przekraczającym 35° nie tworzy się gleba, nie ma roślinności, piargi powstają z materiału sypkiego.
Inne czynniki fizyczne
Inne czynniki fizyczne otaczające żywe organizmy na Ziemi to głównie: elektryczność atmosferyczna, ogień, hałas, ziemskie pole magnetyczne, promieniowanie jonizujące.
elektryczność atmosferyczna działa na organizmy żywe poprzez wyładowania i jonizację powietrza. Na przykład znany jest destrukcyjny efekt uderzenia pioruna w duże drzewa i zwierzęta. Istnieją pewne wzorce częstotliwości uszkodzeń różnych gatunków drzew przez pioruny. Wiąże się to zarówno z kształtem korony, jak iz właściwościami przewodzenia prądu przez korę, np. z szybkością jej zwilżania. Zgodnie z częstotliwością uderzeń piorunów w pierwszej kolejności uszkadza się świerk i sosna, znacznie rzadziej uszkadza brzoza i osika. Piorun powoduje mechaniczne uszkodzenia drzew (rozłupane pnie, pęknięcia), utratę dużych drzew, wpływając tym samym na strukturę drzewostanu, często powodując pożary. Około 21% pożarów lasów w Rosji powstaje z powodu wyładowań atmosferycznych, podczas burz.
Rola atmosferycznych wyładowań elektrycznych polega również na tym, że podczas burzy syntetyzują one z atmosferycznego azotu i tlenu tlenki azotu, które dostają się do gleby wraz z wodą opadową i gromadzą w niej od 4 do 10 kg rocznie na 1 ha w postaci azotanów i kwas azotowy .
Wpływ jonizacji powietrza na ludzi, zwierzęta i rośliny nie został jeszcze dostatecznie zbadany. Jednocześnie niezawodnie ustalono bezpośredni związek między samopoczuciem człowieka a obecnością jonów światła w powietrzu. Wyraża się opinię, że jonizacja powietrza służy materialnej zdolności niektórych roślin do „przewidywania pogody” (spadek fotosyntezy i oddychania, zamykanie szparek i ustanie transpiracji przed burzą na długo przed spadkiem ciśnienia atmosferycznego). Doświadczalnie udowodniono wpływ słabego prądu na system korzeniowy niektórych roślin. Na przykład w sadzonkach świerka i sosny fitomasa wzrasta o 100-120%. Ustalono, że za pomocą działania ukierunkowanego pola elektrycznego można regulować tempo przemieszczania się substancji wewnątrz drzewa, a w konsekwencji tempo jego wzrostu.
Ogień w życiu roślin i zwierząt - dość rzadki, ale bardzo skuteczny czynnik. Pożary, na przykład w lasach, jak wspomniano wcześniej, mogą wystąpić zarówno w sposób naturalny w wyniku uderzenia pioruna, jak i z winy człowieka, jego działań. Dlatego pożar przypisuje się zarówno naturalnym czynnikom środowiskowym, jak i antropogenicznym.
Poważnymi konsekwencjami są nie tylko ukoronowane pożary lasów, obejmujące cały drzewostan, ale także oddolne, które niszczą runo leśne, runo leśne, dolne gałęzie drzew, a często także system korzeniowy. Zwierzęta umierają. Oprócz szkód bezpośrednio po pożarach, pożary powodują degradację drzewostanu. Wzrost maleje. Osłabione drzewa są bardziej podatne na grzyby, takie jak zgnilizna drewna, które z łatwością przenikają przez „rany ogniowe” i są atakowane przez szkodniki.
Pożary lasów w dużym stopniu zmieniają warunki życia roślin i zwierząt. Podczas pożaru w lasach iglastych temperatura dochodzi do 800-900 ° C, w glebie na głębokości 3,5 cm - do 95 ° C, na głębokości 7 cm - do 70 "C. W suchych lasach ściółka i próchnica glebowa wypala się prawie całkowicie Mineralne cząstki wierzchniej warstwy gleby ulegają spiekaniu Tworzą się grudki lub szklista skorupa, które trudno przepuszczają powietrze, wodę i korzenie Gleba jest silnie zagęszczona Na skutek spalania kwasów organicznych i uwalnianie zasad gwałtownie spada kwasowość gleby, w górnych warstwach odczyn często osiąga silnie zasadowy odczyn Pod wpływem wysokiej temperatury górne warstwy gleby ulegają sterylizacji – zamiera mikroflora glebowa, a w głębszych warstwach zmienia się jej skład, grupy najważniejsze dla życia roślin są zubożone. Tak więc w glebach lasów iglastych po pożarach dominuje aktywność mikroorganizmów powodujących fermentację kwasu masłowego i denitryfikację.
Po pożarach lasów następuje gwałtowna zmiana warunków w zbiorowiskach roślinnych (oświetlenie, zmiany temperatury i inne czynniki mikroklimatyczne), zwłaszcza w przypadku niszczenia drzewostanu, co prowadzi do tego, że w przyszłości spalone tereny zasiedlają gatunki żywych organizmów o różnych cechach adaptacyjnych, które pomagają przetrwać pożar i przetrwać na wzgórzach. Tak więc w roślinach są to głęboko podziemne pąki odnowy, zdolność nasion do pozostawania w glebie przez długi czas i wytrzymywania wysokich temperatur, wytrzymałość na mróz, silne światło itp.
Odnowa na terenach spalonych ma swoje własne cechy. Mchy pionierskie pojawiają się na wypalonych miejscach z zarodników nawianych przez wiatr, po trzech do pięciu lat najliczniejszym z mchów jest „ognisty mech” - Funaria hygrometrica. Z wyższych roślin, Ivan-chai (Chamaenerion angustifolion) szybko zapełnia ogień. Stopniowe zasiedlanie spalonych obszarów występuje również z roślinnością drzewiastą – wierzbą, brzozą, osiką itp. (ryc. 4.26).

Ryż. 4.26. Wpływ ognia na roślinność „kołków” drzew
Trans-Uralski step leśny (według D. F. Fedyunina, 1953):
A - przed ogniem; B - po pożarze; B - rok po pożarze; 1 - wierzba; 2 - brzoza, 3 - osika
Pożary stepowe („oparzenia”) mogą być mniej lub bardziej regularne, związane z działalnością człowieka i odgrywać istotną rolę w życiu organizmów żywych, niekiedy pozytywną dla regulacji wzrostu, odnowy, selekcji gatunkowej i utrzymania stałego składu zielarskiego.
Hałas jako naturalny czynnik środowiskowy dla organizmów żywych jest nieistotny, ale może też mieć istotny wpływ przy nasilonych oddziaływaniach antropogenicznych (hałas generowany podczas eksploatacji pojazdów, wyposażenia przedsiębiorstw przemysłowych i bytowych, instalacji wentylacyjnych i turbin gazowych itp.) .
Wartość ciśnienia akustycznego jest zmieniana i normalizowana w decybelach. Cały zakres dźwięków słyszalnych dla człowieka mieści się w granicach 150 dB. Na naszej planecie życie organizmów toczy się w świecie dźwięków. Na przykład ludzki narząd słuchu jest przystosowany do pewnych stałych lub powtarzających się dźwięków (adaptacja słuchowa). Osoba traci zdolność do pracy bez zwykłych dźwięków. Głośny hałas jest jeszcze bardziej szkodliwy dla ludzkiego zdrowia. Osoby żyjące i pracujące w niekorzystnych warunkach akustycznych wykazują oznaki zmian stanu funkcjonalnego ośrodkowego układu nerwowego i sercowo-naczyniowego.
Badania wykazały wpływ hałasu na organizmy roślinne. W ten sposób rośliny w pobliżu lotnisk, z których stale startują samoloty odrzutowe, doświadczają zahamowania wzrostu, a nawet zanikania poszczególnych gatunków. Szereg prac naukowych wykazało przygnębiające działanie hałasu (około 100 dB przy częstotliwości dźwięku od 31,5 do 90 tys. Hz) na rośliny tytoniu, gdzie stwierdzono zmniejszenie intensywności wzrostu liści, przede wszystkim u młodych roślin. Uwagę naukowców przyciąga również wpływ rytmicznych dźwięków na rośliny. Badania nad wpływem muzyki na rośliny (kukurydza, dynia, petunia, cynia, nagietek), przeprowadzone w 1969 roku przez amerykańskiego muzyka i śpiewaka D. Retolaka, wykazały, że rośliny pozytywnie reagowały na muzykę Bacha i indyjskie melodie muzyczne. Ich pokrój, sucha masa biomasy były najwyższe w porównaniu z kontrolą. A najbardziej zaskakujące jest to, że ich łodygi naprawdę wyciągały się w kierunku źródła tych dźwięków. W tym samym czasie zielone rośliny reagowały na muzykę rockową i ciągłe rytmy bębnów zmniejszeniem wielkości liści i korzeni, zmniejszeniem masy, a wszystkie odeszły od źródła dźwięku, jakby chciały uciec od destrukcyjnego efekt muzyki (ryc. 4.27).

Ryż. 4.27. Rodzaj roślin po akcji różnej muzyki:
A - melodie indyjskie (R. Shankar); B - muzyka J.-S. Kawaler; B - muzyka rockowa (eksperymenty D. Retolak, 1969)
Rośliny, podobnie jak ludzie, reagują na muzykę jako integralny żywy organizm. Według wielu naukowców ich czułymi przewodnikami „nerwowymi” są wiązki łyka, merystem i komórki pobudliwe zlokalizowane w różnych częściach rośliny, połączone ze sobą procesami bioelektrycznymi. Prawdopodobnie ten fakt jest jedną z przyczyn podobieństwa reakcji na muzykę u roślin, zwierząt i ludzi.
Pole magnetyczne Ziemi. Nasza planeta Ziemia ma właściwości magnetyczne. Igła kompasu jest zawsze skierowana wzdłuż południka magnetycznego, wskazując północ na jednym końcu i południe na drugim. Magnetolodzy wykazali, że aby wytworzyć obserwowalne pole geomagnetyczne w centrum Ziemi, konieczne jest umieszczenie gigantycznego cylindrycznego magnesu o średnicy 200 km i długości 4000 km. Oś magnesu ziemskiego znajduje się pod kątem 1,5” do osi obrotu Ziemi, więc bieguny magnetyczne nie pokrywają się z biegunami geograficznymi. Z biegiem czasu bieguny magnetyczne zmieniają swoje położenie. Ustalono, że północny biegun magnetyczny porusza się po powierzchni Ziemi o 20,5 m dziennie, czyli 7,5 km rocznie, a południowy o 30 m (11 km rocznie). jeden biegun i zamknij się przez przestrzeń bliską Ziemi w drugim biegunie. Z powodu tego zjawiska w pobliżu Ziemi powstaje magnetosfera (ryc. 4.28).

Ryż. 4.28. Przekroje południkowe magnetosfery Ziemi:
1 - wiatr słoneczny; 2 - przód szoku; 3 - wnęka magnetyczna; 4 - magnetopauza; 5 - górna granica szczeliny magnetosferycznej; 6 - płaszcz plazmowy; 7 - zewnętrzny pas radiacyjny; 8 - wewnętrzny pas radiacyjny lub plazmasfera; 9 - warstwa neutralna; 10 - warstwa plazmy
Opóźnia przepływ cząstek naładowanych energią słoneczną, zwanych plazmą lub wiatrem słonecznym, uniemożliwiając im dotarcie do powierzchni planety. Wiatr słoneczny niejako krąży wokół Ziemi i przesuwa się na stronę nocną, pociągając z kolei magnetyczne linie siły w tym samym kierunku. Odkształcenie linii pola magnetycznego wynika z faktu, że przepływy plazmy słonecznej niosą ze sobą niejako „zamrożone” pole magnetyczne, które oddziałuje z magnetosferą Ziemi. W ciągu ostatnich 600 tysięcy lat paleomagnetolodzy odnotowali 12 epok odwrócenia pola geomagnetycznego (tabela 4.10).