Genetik kod kodlangan. Genetik kod nima: umumiy ma'lumot
5-ma'ruza. Genetik kod
Kontseptsiyaning ta'rifi
Genetik kod - bu DNKdagi nukleotidlar ketma-ketligidan foydalangan holda oqsillardagi aminokislotalarning ketma-ketligi haqidagi ma'lumotlarni yozib olish tizimi.
DNK oqsil sintezida bevosita ishtirok etmaganligi sababli kod RNK tilida yozilgan. RNK tarkibida timin o'rniga urasil mavjud.
Genetik kodning xususiyatlari
1. Uchlik
Har bir aminokislota 3 ta nukleotidlar ketma-ketligi bilan kodlangan.
Ta'rif: triplet yoki kodon - bu bitta aminokislotalarni kodlaydigan uchta nukleotidlar ketma-ketligi.
Kod monoplet bo'lishi mumkin emas, chunki 4 (DNKdagi turli nukleotidlar soni) 20 dan kam. Kod dublet bo'lishi mumkin emas, chunki 16 (2 dan 4 ta nukleotidning birikmalari va almashinishlari soni) 20 dan kam. Kod uchlik bo'lishi mumkin, chunki 64 (4 dan 3 gacha kombinatsiyalar va almashtirishlar soni) 20 dan ortiq.
2. Degeneratsiya.
Barcha aminokislotalar, metionin va triptofandan tashqari, bir nechta tripletlar bilan kodlangan:
1 uchlik uchun 2 AK = 2.
9 AK, har biri 2 ta uchlik = 18.
1 AK 3 uchlik = 3.
4 ta uchlikdan 5 AK = 20.
6 ta uchlikdan 3 ta AK = 18.
Jami 61 triplet 20 ta aminokislotalarni kodlaydi.
3. Genlararo tinish belgilarining mavjudligi.
Ta'rif:
Gen - bitta polipeptid zanjiri yoki bitta molekulani kodlaydigan DNK bo'limi tRNK, rRNK yokisRNK.
GenlartRNK, rRNK, sRNKoqsillar kodlanmagan.
Har bir polipeptid-kodlovchi genning oxirida RNK to'xtash kodonlarini yoki to'xtash signallarini kodlovchi 3 ta uchlikdan kamida bittasi mavjud. mRNKda ular quyidagi shaklga ega: UAA, UAG, UGA . Ular eshittirishni tugatadilar (tugaydilar).
An'anaviy ravishda kodon ham tinish belgilariga tegishli AVG - yetakchilar ketma-ketligidan keyingi birinchi. (8-ma'ruzaga qarang) U bosh harf vazifasini bajaradi. Bu holatda u formilmetioninni (prokaryotlarda) kodlaydi.
4. Aniqlik.
Har bir triplet faqat bitta aminokislotani kodlaydi yoki tarjima terminatoridir.
Istisno - bu kodon AVG . Prokaryotlarda birinchi holatda (katta harf) formilmetioninni, boshqa har qanday holatda esa metioninni kodlaydi.
5. Kompaktlik, yoki intragenik tinish belgilarining yo'qligi.
Gen ichida har bir nukleotid muhim kodonning bir qismidir.
1961 yilda Seymur Benzer va Frensis Krik kodning uchlik tabiatini va uning ixchamligini eksperimental tarzda isbotladilar.
Tajribaning mohiyati: "+" mutatsiyasi - bitta nukleotidning kiritilishi. "-" mutatsiyasi - bitta nukleotidni yo'qotish. Genning boshida bitta "+" yoki "-" mutatsiya butun genni buzadi. Ikki tomonlama "+" yoki "-" mutatsiya ham butun genni buzadi.
Genning boshida uch marta "+" yoki "-" mutatsiya uning faqat bir qismini buzadi. To'rt martalik "+" yoki "-" mutatsiya yana butun genni buzadi.
Tajriba buni isbotlaydi Kod transkripsiya qilinadi va gen ichida tinish belgilari yo'q. Tajriba ikkita qo'shni fag genida o'tkazildi va bundan tashqari, genlar orasidagi tinish belgilarining mavjudligi.
6. Ko'p qirralilik.
Er yuzida yashovchi barcha mavjudotlar uchun genetik kod bir xil.
1979 yilda Burrell ochildi ideal inson mitoxondriya kodi.
Ta'rif:
Bu "ideal" deb ataladi genetik kod, bunda kvazi-dubl kodning degeneratsiya qoidasi qanoatlansa: Agar ikkita tripletda birinchi ikkita nukleotid mos tushsa va uchinchi nukleotidlar bir sinfga tegishli bo'lsa (ikkalasi ham purinlar yoki ikkalasi ham pirimidinlar), u holda bu tripletlar bir xil aminokislota.
Universal kodda bu qoidadan ikkita istisno mavjud. Umumjahondagi ideal koddan ikkala og'ish ham asosiy nuqtalarga tegishli: oqsil sintezining boshlanishi va oxiri:
Kodon | Universal kod | Mitoxondriyal kodlar |
|||
Umurtqali hayvonlar | Umurtqasizlar | Xamirturush | O'simliklar |
||
STOP | STOP |
||||
UA bilan | |||||
A G A | STOP | ||||
STOP | 230 ta almashtirish kodlangan aminokislota sinfini o'zgartirmaydi. yirtiluvchanlikka. 1956 yilda Georgiy Gamov bir-biriga o'xshash kodning variantini taklif qildi. Gamow kodiga ko'ra, gendagi uchinchidan boshlab har bir nukleotid 3 ta kodonning bir qismidir. Genetik kod dekodlanganda, u bir-biriga mos kelmasligi ma'lum bo'ldi, ya'ni. Har bir nukleotid faqat bitta kodonning bir qismidir. Bir-biriga mos keladigan genetik kodning afzalliklari: ixchamlik, oqsil strukturasining nukleotidni kiritish yoki yo'q qilishga kamroq bog'liqligi. Kamchilik: oqsil tuzilishi nukleotidlarni almashtirishga va qo'shnilarga cheklovlarga juda bog'liq. 1976 yilda phX174 fagining DNKsi ketma-ketlashtirildi. U 5375 nukleotiddan tashkil topgan bir zanjirli dumaloq DNKga ega. Fag 9 ta oqsilni kodlashi ma'lum edi. Ulardan 6 tasi uchun birin-ketin joylashgan genlar aniqlangan. Ma'lum bo'lishicha, bir-biriga o'xshashlik bor. Gen E butunlay gen ichida joylashgan D . Uning boshlang'ich kodoni bitta nukleotidning ramka siljishidan kelib chiqadi. Gen J gen tugagan joydan boshlanadi D . Genning kodoni boshlang J genning to'xtash-kodon bilan ustma-ust tushadi D ikkita nukleotidning siljishi natijasida. Qurilish uchga ko'paytirilmagan nukleotidlar soni tomonidan "o'qish ramkasining siljishi" deb ataladi. Bugungi kunga kelib, bir-biriga o'xshashlik faqat bir nechta faglar uchun ko'rsatilgan. DNKning axborot sig'imi Yer yuzida 6 milliard odam yashaydi. Ular haqida irsiy ma'lumotlar 4x10 13 kitob sahifalari. Ushbu sahifalar 6 ta NSU binosining maydonini egallaydi. 6x10 9 spermatozoidlar yarim oyukni egallaydi. Ularning DNKsi to'rtdan bir qismidan kamroq joyni egallaydi. | ||||
Kimyoviy tarkibi va DNK molekulasining strukturaviy tashkil etilishi.
Molekulalar nuklein kislotalar ko'p yuzlab va hatto millionlab nukleotidlardan tashkil topgan juda uzun zanjirlardir. Har qanday nuklein kislota faqat to'rt turdagi nukleotidlarni o'z ichiga oladi. Nuklein kislota molekulalarining funktsiyalari ularning tuzilishiga, tarkibidagi nukleotidlarga, ularning zanjirdagi soniga va molekuladagi birikmaning ketma-ketligiga bog'liq.
Har bir nukleotid uchta komponentdan iborat: azotli asos, uglevod va fosfor kislotasi. IN birikma har bir nukleotid DNK azotli asoslarning to'rt turidan birini (adenin - A, timin - T, guanin - G yoki sitozin - C), shuningdek dezoksiriboza uglerod va fosfor kislotasi qoldig'ini o'z ichiga oladi.
Shunday qilib, DNK nukleotidlari faqat azotli asosning turiga ko'ra farqlanadi.
DNK molekulasi ma'lum bir ketma-ketlikda zanjirda bog'langan juda ko'p nukleotidlardan iborat. DNK molekulasining har bir turi o'z nukleotidlari soni va ketma-ketligiga ega.
DNK molekulalari juda uzun. Masalan, insonning bir hujayrasidan (46 xromosoma) DNK molekulalaridagi nukleotidlar ketma-ketligini harflar bilan yozish uchun taxminan 820 000 sahifali kitob kerak bo'ladi. Muqobil to'rt turdagi nukleotidlar hosil bo'lishi mumkin cheksiz to'plam DNK molekulalarining variantlari. DNK molekulalarining bu strukturaviy xususiyatlari ularga organizmlarning barcha xususiyatlari haqida juda katta hajmdagi ma'lumotlarni saqlashga imkon beradi.
1953 yilda amerikalik biolog J. Uotson va ingliz fizigi F. Krik DNK molekulasining tuzilishi modelini yaratdilar. Olimlar har bir DNK molekulasi o‘zaro bog‘langan va spiral tarzda o‘ralgan ikkita zanjirdan iborat ekanligini aniqladilar. Bu ikki tomonlama spiralga o'xshaydi. Har bir zanjirda to'rt turdagi nukleotidlar ma'lum bir ketma-ketlikda almashadilar.
Nukleotid DNK tarkibi orasida farqlanadi har xil turlari bakteriyalar, qo'ziqorinlar, o'simliklar, hayvonlar. Ammo u yoshga qarab o'zgarmaydi, o'zgarishlarga ozgina bog'liq muhit. Nukleotidlar juftlashgan, ya'ni har qanday DNK molekulasidagi adenin nukleotidlari soni timidin nukleotidlari soniga (A-T), sitozin nukleotidlari soni esa guanin nukleotidlari soniga (C-G) teng bo'ladi. Buning sababi, DNK molekulasida ikkita zanjirning bir-biriga bog'lanishi ma'lum bir qoidaga bo'ysunadi, ya'ni: bir zanjirning adenini har doim ikkita vodorod aloqasi bilan faqat boshqa zanjirning Timin bilan bog'lanadi va guanin - sitozin bilan uchta vodorod aloqasi orqali, ya'ni bir molekula DNKning nukleotid zanjirlari bir-birini to'ldiruvchi, bir-birini to'ldiradi.
Nuklein kislota molekulalari - DNK va RNK - nukleotidlardan tashkil topgan. DNK nukleotidlariga azotli asos (A, T, G, C), uglevod dezoksiriboza va fosfor kislotasi molekulasi qoldig'i kiradi. DNK molekulasi ikki tomonlama spiral, to'ldiruvchilik printsipiga ko'ra vodorod aloqalari bilan bog'langan ikkita zanjirdan iborat. DNKning vazifasi irsiy ma'lumotlarni saqlashdir.
DNKning xossalari va vazifalari.
DNK tashuvchi hisoblanadi genetik ma'lumot, genetik kod yordamida nukleotidlar ketma-ketligi sifatida qayd etilgan. DNK molekulalari ikkita asosiy bilan bog'langan tirik mavjudotlarning xususiyatlari organizmlar - irsiyat va o'zgaruvchanlik. DNK replikatsiyasi deb ataladigan jarayon davomida asl zanjirning ikkita nusxasi hosil bo'ladi, ular bo'linganda qiz hujayralar tomonidan meros qilib olinadi, natijada hosil bo'lgan hujayralar genetik jihatdan asl nusxaga o'xshash bo'ladi.
Genetik ma'lumotlar transkripsiya (RNK molekulalarining DNK shablonida sintezi) va translyatsiya (RNK shablonidagi oqsillarni sintezi) jarayonlarida gen ekspressiyasi paytida amalga oshiriladi.
Nukleotidlar ketma-ketligi haqidagi ma'lumotlarni "kodlaydi" har xil turlari RNK: axborot yoki matritsa (mRNK), ribosoma (rRNK) va transport (tRNK). Ushbu turdagi RNKlarning barchasi transkripsiya jarayonida DNKdan sintezlanadi. Ularning oqsil biosintezidagi roli (tarjima jarayoni) boshqacha. Messenger RNK oqsildagi aminokislotalarning ketma-ketligi haqidagi ma'lumotlarni o'z ichiga oladi, ribosoma RNK ribosomalar uchun asos bo'lib xizmat qiladi (asosiy vazifasi mRNK asosida individual aminokislotalardan oqsillarni yig'ish bo'lgan murakkab nukleoprotein komplekslari), transfer RNKlari aminokislotalarni etkazib beradi. kislotalar oqsil yig'ilish joyiga - ribosomaning faol markaziga, mRNKda "emaklashuvchi".
Genetik kod, uning xossalari.
Genetik kod- nukleotidlar ketma-ketligi yordamida oqsillarning aminokislotalar ketma-ketligini kodlashning barcha tirik organizmlarga xos bo'lgan usuli. XUSUSIYATLARI:
- Uchlik- kodning mazmunli birligi uchta nukleotidning (uchlik yoki kodon) birikmasidir.
- Davomiylik- uchlik o'rtasida tinish belgilari yo'q, ya'ni ma'lumotlar uzluksiz o'qiladi.
- Bir-biriga mos kelmaslik- bir xil nukleotid bir vaqtning o'zida ikki yoki undan ortiq tripletlarning bir qismi bo'lishi mumkin emas (bir nechta ramka almashinuvi oqsillarini kodlaydigan viruslar, mitoxondriyalar va bakteriyalarning bir-biriga o'xshash ba'zi genlari uchun kuzatilmaydi).
- O'ziga xoslik (o'ziga xoslik)- ma'lum bir kodon faqat bitta aminokislotaga to'g'ri keladi (ammo UGA kodonida mavjud Euplotes crassus ikkita aminokislotalarni kodlaydi - sistein va selenosistein)
- Degeneratsiya (ortiqchalik)- bir aminokislotaga bir nechta kodon mos kelishi mumkin.
- Ko'p qirralilik- genetik kod turli darajadagi murakkablikdagi organizmlarda bir xil ishlaydi - viruslardan odamlargacha (usullar bunga asoslanadi) genetik muhandislik; bir qator istisnolar mavjud, ular quyidagi standart genetik kod bo'limidagi Variatsiyalar jadvalida ko'rsatilgan).
- Shovqinga qarshi immunitet- kodlangan aminokislotalar sinfining o'zgarishiga olib kelmaydigan nukleotid almashtirish mutatsiyalari deyiladi. konservativ; kodlangan aminokislota sinfining o'zgarishiga olib keladigan nukleotidlarni almashtirish mutatsiyalari deyiladi radikal.
5. DNKning avtoko'payishi. Replikon va uning faoliyati .
Meros bilan birga bo'lgan nuklein kislota molekulalarining o'z-o'zini ko'paytirish jarayoni (hujayradan hujayraga) aniq nusxalari genetik ma'lumotlar; R. o'ziga xos fermentlar to'plami ishtirokida amalga oshiriladi (helikaz<spiral>molekulaning yechilishini nazorat qilish DNK, DNK-polimeraza<DNK polimeraza> I va III, DNK-ligaza<DNK ligaza>), replikatsiya vilkasini hosil qilish bilan yarim konservativ tarzda davom etadi<replikatsiya vilkasi>; sxemalardan birida<yetakchi qator> to'ldiruvchi zanjirning sintezi uzluksiz va boshqa tomondan<orqada qolgan ip> Dkazaki fragmentlarining hosil bo'lishi tufayli yuzaga keladi<Okazaki parchalari>; R. - xato darajasi 10 -9 dan oshmaydigan yuqori aniqlikdagi jarayon; eukariotlarda R. bir vaqtning o'zida bir molekulaning bir nechta nuqtasida paydo bo'lishi mumkin DNK; tezlik R. eukariotlarda sekundiga 100 ga yaqin, bakteriyalarda esa 1000 ga yaqin nukleotidlar mavjud.
6. Eukaryotik genomning tashkiliy darajalari .
Eukaryotik organizmlarda transkripsiyani tartibga solish mexanizmi ancha murakkab. Eukaryotik genlarni klonlash va sekvensiyalash natijasida transkripsiya va translatsiyada ishtirok etuvchi maxsus ketma-ketliklar topildi.
Eukaryotik hujayra quyidagi belgilar bilan tavsiflanadi:
1. DNK molekulasida intron va ekzonlarning mavjudligi.
2. mRNKning yetilishi - intronlarning kesilishi va ekzonlarning tikilishi.
3. Transkripsiyani tartibga soluvchi tartibga soluvchi elementlarning mavjudligi, masalan: a) promotorlar - har birida o'ziga xos polimeraza egallagan 3 xil. Pol I ribosoma genlarini, Pol II oqsil strukturaviy genlarini, Pol III kichik RNKlarni kodlovchi genlarni replikatsiya qiladi. Pol I va Pol II promotori transkripsiyani boshlash joyi oldida, Pol III promouteri strukturaviy gen ichida joylashgan; b) modulyatorlar - transkripsiya darajasini kuchaytiruvchi DNK ketma-ketliklari; v) kuchaytirgichlar - transkripsiya darajasini oshiradigan va genning kodlash qismiga nisbatan pozitsiyasidan va RNK sintezining boshlang'ich nuqtasi holatidan qat'iy nazar harakat qiladigan ketma-ketliklar; d) terminatorlar - tarjimani ham, transkripsiyani ham to'xtatuvchi o'ziga xos ketma-ketliklar.
Bu ketma-ketliklar boshlang'ich kodonga nisbatan birlamchi tuzilishi va joylashuvi bilan prokaryotik ketma-ketliklardan farq qiladi va bakterial RNK polimeraza ularni "tanimaydi". Shunday qilib, prokaryotik hujayralardagi eukaryotik genlarni ifodalash uchun genlar prokaryotik tartibga soluvchi elementlarning nazorati ostida bo'lishi kerak. Bu holat ifoda vektorlarini qurishda hisobga olinishi kerak.
7. Xromosomalarning kimyoviy va strukturaviy tarkibi .
Kimyoviy xromosoma tarkibi - DNK - 40%, giston oqsillari - 40%. Giston bo'lmagan - 20% bir oz RNK. Lipidlar, polisaxaridlar, metall ionlari.
Xromosomaning kimyoviy tarkibi oqsillar, uglevodlar, lipidlar va metallar bilan nuklein kislotalar majmuasidir. Xromosoma gen faolligini tartibga soladi va kimyoviy yoki radiatsiyaviy zararlanganda uni tiklaydi.
Strukturaviy????
Xromosomalar- nukleoprotein strukturaviy elementlar organizmning irsiy ma'lumotlarini o'z ichiga olgan DNKni o'z ichiga olgan hujayra yadrolari o'z-o'zini ko'paytirishga qodir, tarkibiy va funktsional individuallikka ega va uni bir necha avlodlar davomida saqlab qoladi.
mitotik siklda xromosomalarning strukturaviy tashkil etilishining quyidagi xususiyatlari kuzatiladi:
Xromosomalarning strukturaviy tashkil etilishining mitotik va interfaza shakllari mavjud bo'lib, ular mitotik siklda o'zaro bir-biriga aylanadi - bular funktsional va fiziologik o'zgarishlardir.
8. Eukariotlarda irsiy materialning qadoqlanish darajalari .
Eukariotlarning irsiy materialini tashkil qilishning strukturaviy va funksional darajalari
Irsiyat va o'zgaruvchanlik quyidagilarni ta'minlaydi:
1) individual (diskret) meros va individual xususiyatlarning o'zgarishi;
2) ma'lum bir biologik tur organizmlarining morfofunksional xususiyatlarining butun majmuasini har bir avlodning individlarida ko'paytirish;
3) irsiy moyilliklarni ko'paytirish jarayonida jinsiy ko'payish bilan turlarda qayta taqsimlash, buning natijasida avlod ota-onalardagi kombinatsiyasidan farq qiladigan xususiyatlarning kombinatsiyasiga ega. Belgilarning irsiyat va o'zgaruvchanligi va ularning to'plamlari genetik materialning strukturaviy va funktsional tashkil etilishi tamoyillaridan kelib chiqadi.
Eukaryotik organizmlarning irsiy materialini tashkil qilishning uchta darajasi mavjud: gen, xromosoma va genomik (genotip darajasi).
Elementar tuzilma gen darajasi gen sifatida xizmat qiladi. Genlarning ota-onadan naslga o'tishi ma'lum xususiyatlarning rivojlanishi uchun zarurdir. Biologik o'zgaruvchanlikning bir nechta shakllari ma'lum bo'lsa-da, faqat genlar tuzilishining buzilishi irsiy ma'lumotlarning ma'nosini o'zgartiradi, unga muvofiq o'ziga xos xususiyatlar va xususiyatlar shakllanadi. Gen darajasining mavjudligi tufayli individual, alohida (diskret) va mustaqil meros va individual xususiyatlarning o'zgarishi mumkin.
Eukaryotik hujayralardagi genlar xromosomalar bo'ylab guruhlarga bo'linadi. Bu hujayra yadrosining tuzilmalari bo'lib, ular individuallik va avlodlar davomida individual strukturaviy xususiyatlarni saqlab qolish bilan o'zlarini ko'paytirish qobiliyati bilan ajralib turadi. Xromosomalarning mavjudligi irsiy materialni tashkil etishning xromosoma darajasini aniqlashni belgilaydi. Genlarning xromosomalarga joylashishi belgilarning nisbiy merosiga ta'sir qiladi va gen funktsiyasiga uning bevosita genetik muhiti - qo'shni genlar ta'sir qilishiga imkon beradi. Irsiy materialning xromosoma tuzilishi xizmat qiladi zaruriy shart jinsiy ko'payish paytida ota-onalarning nasl-nasabidagi irsiy moyilligini qayta taqsimlash.
Turli xil xromosomalar bo'yicha taqsimlanishiga qaramay, genlarning butun to'plami funktsional ravishda o'zini bir butun sifatida shakllantiradi. yagona tizim, irsiy materialni tashkil etishning genomik (genotipik) darajasini ifodalaydi. Bu darajada bitta va turli xromosomalarda lokalizatsiya qilingan irsiy moyilliklarning keng o'zaro ta'siri va o'zaro ta'siri mavjud. Natijada turli xil irsiy moyilliklarning genetik ma'lumotlarining o'zaro muvofiqligi va natijada ontogenez jarayonida vaqt, joy va intensivlik bo'yicha muvozanatlangan belgilarning rivojlanishi. Genlarning funksional faolligi, replikatsiya usuli va irsiy materialdagi mutatsion o'zgarishlar ham organizm yoki umuman hujayra genotipining xususiyatlariga bog'liq. Buni, masalan, hukmronlik xususiyatining nisbiyligi tasdiqlaydi.
Eu - va geterokromatin.
Hujayra bo'linishi paytida ba'zi xromosomalar kondensatsiyalangan va kuchli rangli ko'rinadi. Bunday farqlar geteropiknoz deb ataldi. Atama " heteroxromatin" Euxromatin mavjud - mitoz xromosomalarning asosiy qismi bo'lib, ular mitoz paytida odatiy siqilish va dekompaktsiya siklini boshdan kechiradi va heteroxromatin- xromosomalarning doimo ixcham holatda bo'lgan hududlari.
Eukariotlarning aksariyat turlarida xromosomalar ikkalasini ham o'z ichiga oladi ew- va geteroxromatik hududlar, ikkinchisi genomning muhim qismini tashkil qiladi. Geterokromatin perisentromerik, ba'zan peritomerik hududlarda joylashgan. Xromosomalarning evromatik qo'llarida geteroxromatik hududlar topilgan. Ular heteroxromatinning evromatin tarkibiga kiritilishi (interkalatsiyasi) kabi ko'rinadi. Bunday heteroxromatin interkalyar deb ataladi. Xromatinning siqilishi. Evromatin va heteroxromatin siqilish davrlarida farqlanadi. Euhr. interfazadan interfazaga, hetero siqilish-dekompaktsiyaning to'liq siklidan o'tadi. nisbatan ixchamlik holatini saqlaydi. Differensial barqarorlik. Geteroxromatinning turli joylari turli bo'yoqlar bilan bo'yalgan, ba'zi joylari bitta, boshqalari bir nechta. Turli bo'yoqlardan foydalanish va geteroxromatik hududlarni buzadigan xromosomalarni qayta tashkil etish yordamida Drosophiladagi dog'larga yaqinlik qo'shni mintaqalardan farq qiladigan ko'plab kichik hududlarni tavsiflash mumkin bo'ldi.
10. Metafaza xromosomasining morfologik xususiyatlari .
Metafaza xromosomasi birlamchi siqilish mintaqasida - sentromerada bir-biriga bog'langan dezoksiribonukleoprotein - xromatidlarning ikki bo'ylama ipidan iborat. Centromere - maxsus tarzda tashkil etilgan sayt ikkala opa-singil xromatidlar uchun umumiy xromosomalar. Tsentromera xromosoma tanasini ikki qismga ajratadi. Birlamchi siqilish joyiga qarab, mavjud quyidagi turlari xromosomalar: teng qo'llar (metasentrik), sentromera o'rtada joylashganda va qo'llar taxminan teng uzunlik; teng bo'lmagan qo'llar (submetasentrik), sentromera xromosomaning o'rtasidan siljiganida va qo'llar teng bo'lmagan uzunlikda; tayoqchali (akrosentrik), sentromera xromosomaning bir uchiga siljiganida va bir qo'li juda qisqa bo'ladi. Nuqtali (telotsentrik) xromosomalar ham bor, ularda bitta qo'l yo'q, lekin ular inson karyotipida (xromosomalar to'plami) mavjud emas; Ba'zi xromosomalarda yo'ldosh deb ataladigan hududni xromosoma tanasidan ajratib turadigan ikkilamchi siqilishlar bo'lishi mumkin.
GENETIK KOD - DNK molekulalarida (ba'zi viruslarda - RNK) nukleotid asoslari ketma-ketligi ko'rinishidagi irsiy ma'lumotni qayd qilish tizimi, oqsil molekulalarida (polipeptidlar) birlamchi strukturani (aminokislota qoldiqlarining joylashishini) aniqlaydi. Genetik kod muammosi DNKning genetik rolini isbotlagandan so'ng (amerikalik mikrobiologlar O. Averi, K. Makleod, M. Makkarti, 1944) va uning tuzilishini dekodlashdan so'ng (J. Uotson, F. Krik, 1953) shakllantirildi. genlar fermentlarning tuzilishi va funktsiyalarini belgilaydi (J. Beadle va E. Tatem, 1941 "bitta gen - bitta ferment" tamoyili) va oqsilning fazoviy tuzilishi va faolligi uning birlamchi tuzilishiga bog'liqligi borligi. (F. Sanger, 1955). 4 ta nuklein kislota asoslarining birikmalari polipeptidlardagi 20 ta umumiy aminokislotalar qoldiqlarining almashinishini qanday aniqlaydi, degan savol birinchi marta 1954 yilda G. Gamov tomonidan qoʻyilgan.
F. Krik va boshqa olimlar 1961 yilda T4 bakteriofagining genlaridan birida bir juft nukleotidning qo'shilishi va yo'q qilinishining o'zaro ta'sirini o'rgangan tajribaga asoslanib, F. Krik va boshqa olimlar umumiy xususiyatlar genetik kod: triplet, ya'ni polipeptid zanjiridagi har bir aminokislota qoldig'i gen DNKsidagi uchta asos (triplet yoki kodon) to'plamiga to'g'ri keladi; gen ichidagi kodonlar belgilangan nuqtadan, bir yo'nalishda va "vergullarsiz" o'qiladi, ya'ni kodonlar bir-biridan hech qanday belgilar bilan ajratilmaydi; degeneratsiya yoki ortiqcha - bir xil aminokislota qoldig'i bir nechta kodonlar (sinonim kodonlar) bilan kodlanishi mumkin. Mualliflar kodonlar bir-birining ustiga chiqmaydi, deb taxmin qilishgan (har bir asos faqat bitta kodonga tegishli). Sintetik messenjer RNK (mRNK) nazorati ostida hujayrasiz oqsil sintezi tizimi yordamida tripletlarning kodlash qobiliyatini bevosita o'rganish davom ettirildi. 1965 yilga kelib, genetik kod S. Ochoa, M. Nirenberg va X. G. Korana asarlarida to'liq deşifrlangan. Genetik kod sirlarini ochish XX asrda biologiyaning ajoyib yutuqlaridan biri edi.
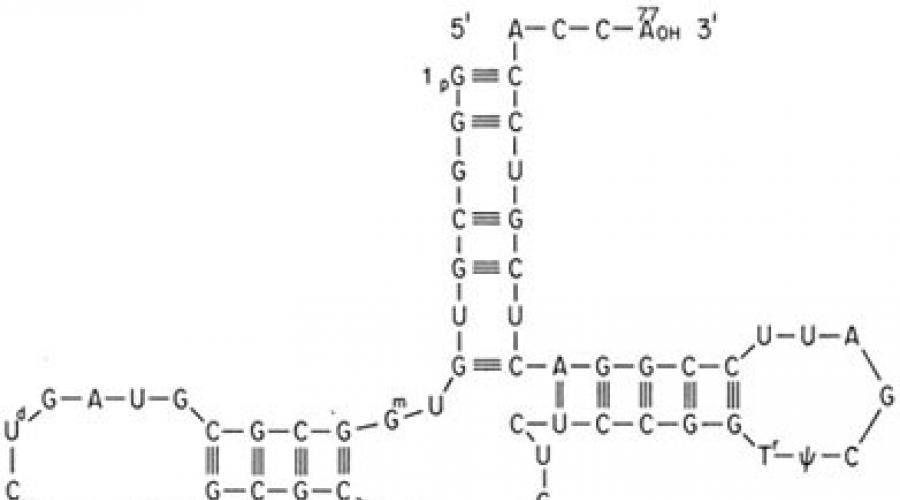
Hujayrada genetik kodning amalga oshirilishi ikkita matritsa jarayoni - transkripsiya va tarjima jarayonida sodir bo'ladi. Gen va oqsil o'rtasidagi vositachi mRNK bo'lib, u DNK zanjirlaridan birida transkripsiya paytida hosil bo'ladi. Bunday holda, oqsilning birlamchi tuzilishi haqida ma'lumotni olib yuruvchi DNK asoslari ketma-ketligi mRNK asoslari ketma-ketligi shaklida "qayta yoziladi". Keyin, ribosomalarga tarjima paytida, mRNKning nukleotidlar ketma-ketligi transfer RNKlari (tRNK) tomonidan o'qiladi. Ikkinchisida aminokislota qoldig'i biriktirilgan qabul qiluvchi uchi va mos keladigan mRNK kodonini taniydigan adapter uchi yoki antikodon tripleti mavjud. Kodon va antikodonning o'zaro ta'siri komplementar asoslar juftligi asosida sodir bo'ladi: Adenin (A) - Uratsil (U), Guanin (G) - Sitozin (C); bu holda mRNKning asosiy ketma-ketligi sintezlangan oqsilning aminokislotalar ketma-ketligiga aylantiriladi. Turli organizmlar Ular bir xil aminokislota uchun turli chastotali turli xil sinonim kodonlardan foydalanadilar. Polipeptid zanjirini kodlovchi mRNKni o'qish metionin aminokislotalariga mos keladigan AUG kodonidan boshlanadi (boshlanadi). Kamroq, prokaryotlarda boshlang'ich kodonlar GUG (valin), UUG (leytsin), AUU (izoleysin) va eukariotlarda - UUG (leytsin), AUA (izoleysin), ACG (treonin), CUG (leytsin). Bu tarjima paytida o'qishning ramkasi yoki fazasini belgilaydi, ya'ni mRNKning butun nukleotidlar ketma-ketligi tRNKning tripleti bo'yicha uch marta o'qiladi, ko'pincha to'xtash kodonlari deb ataladigan uchta terminator kodonlaridan birortasi uchratilguncha o'qiladi. mRNK: UAA, UAG, UGA (jadval). Ushbu tripletlarni o'qish polipeptid zanjirining sintezini yakunlashga olib keladi.
AUG va stop kodonlari mos ravishda polipeptidlarni kodlovchi mRNK mintaqalarining boshida va oxirida paydo bo'ladi.
Genetik kod deyarli universaldir. Bu shuni anglatadiki, ba'zi kodonlarning ma'nosida ozgina farqlar mavjud turli ob'ektlar, va bu, birinchi navbatda, muhim bo'lishi mumkin bo'lgan terminator kodonlariga taalluqlidir; masalan, ba'zi eukariotlar va mikoplazmalarning mitoxondriyalarida UGA triptofanni kodlaydi. Bundan tashqari, bakteriyalar va eukariotlarning ba'zi mRNKlarida UGA g'ayrioddiy aminokislota - selenotsisteinni va arxebakteriyalardan birida UAG - pirrolizini kodlaydi.
Genetik kod tasodifan paydo bo'lgan nuqtai nazar mavjud ("muzlatilgan imkoniyat" gipotezasi). Bu rivojlangan bo'lishi ehtimoli ko'proq. Aminokislota uchta asosdan faqat ikkitasi bilan aniqlanganda, mitoxondriyada "uchdan ikkitasi" qoidasiga ko'ra o'qiladigan kodning oddiyroq va, ehtimol, qadimiy versiyasi mavjudligi bu taxminni tasdiqlaydi. uchlikda.
Lit.: Krik F. N. a. O. Oqsillar uchun genetik kodning umumiy tabiati // Tabiat. 1961. jild. 192; Genetik kod. N.Y., 1966 yil; Ichas M. Biologik kod. M., 1971; Inge-Vechtomov S.G. Genetik kod qanday o'qiladi: qoidalar va istisnolar // Zamonaviy tabiatshunoslik. M., 2000. T. 8; Ratner V. A. Genetik kod tizim sifatida // Soros ta'lim jurnali. 2000. T. 6. No 3.
S. G. Inge-Vechtomov.
Genetik kod - bu DNK yoki RNKdagi nukleotidlar ketma-ketliklarining ma'lum bir almashinishiga asoslangan, oqsildagi aminokislotalarga mos keladigan kodonlarni hosil qiluvchi nuklein kislota molekulalarida irsiy ma'lumotni qayd qilish tizimi.
Genetik kodning xususiyatlari.
Genetik kod bir nechta xususiyatlarga ega.
Uchlik.
Degeneratsiya yoki ortiqchalik.
Aniqlik.
Polarlik.
Bir-biriga mos kelmaslik.
Kompaktlik.
Ko'p qirralilik.
Shuni ta'kidlash kerakki, ba'zi mualliflar kodning boshqa xususiyatlarini ham taklif qilishadi kimyoviy xususiyatlar nukleotidlar kodiga kiritilgan yoki organizm oqsillarida alohida aminokislotalarning paydo bo'lish chastotasi va boshqalar. Biroq, bu xususiyatlar yuqorida sanab o'tilganlardan kelib chiqadi, shuning uchun biz ularni o'sha erda ko'rib chiqamiz.
A. Uchlik. Genetik kod, ko'p narsalar kabi, murakkab uyushgan tizim eng kichik strukturaviy va eng kichik funktsional birlikka ega. Triplet genetik kodning eng kichik tarkibiy birligidir. U uchta nukleotiddan iborat. Kodon - eng kichik funktsional birlik genetik kod. Odatda, mRNKning tripletlari kodonlar deb ataladi. Genetik kodda kodon bir nechta funktsiyalarni bajaradi. Birinchidan, uning asosiy vazifasi bitta aminokislotani kodlashidir. Ikkinchidan, kodon aminokislotalarni kodlamasligi mumkin, ammo bu holda u boshqa funktsiyani bajaradi (pastga qarang). Ta'rifdan ko'rinib turibdiki, triplet xarakterlovchi tushunchadir boshlang'ich strukturaviy birlik genetik kod (uch nukleotid). Kodon - xarakterlaydi elementar semantik birlik genom - uchta nukleotid bitta aminokislotaning polipeptid zanjiriga biriktirilishini aniqlaydi.
Elementar strukturaviy birlik dastlab nazariy jihatdan shifrlangan, keyin esa uning mavjudligi eksperimental tarzda tasdiqlangan. Haqiqatan ham, 20 ta aminokislotalarni bir yoki ikkita nukleotid bilan kodlash mumkin emas, chunki to'rtta nukleotiddan faqat 4 tasi mavjud bo'lib, bu 4 3 = 64 ta variantni beradi, bu tirik organizmlarda mavjud bo'lgan aminokislotalar sonini qoplaydi (1-jadvalga qarang).
Jadvalda keltirilgan 64 ta nukleotid birikmasi ikkita xususiyatga ega. Birinchidan, 64 ta triplet variantlardan faqat 61 tasi kodon va har qanday aminokislotalarni kodlaydi, ular deyiladi. sezuvchi kodonlar. Uchta uchlik kodlanmaydi
aminokislotalar a - translatsiyaning tugashini ko'rsatuvchi to'xtash signallari. Bunday uchta uchlik bor - UAA, UAG, UGA, ular "ma'nosiz" (bema'nilik kodonlari) deb ham ataladi. Tripletdagi bir nukleotidni boshqasi bilan almashtirish bilan bog'liq bo'lgan mutatsiya natijasida his kodonidan ma'nosiz kodon paydo bo'lishi mumkin. Ushbu turdagi mutatsiya deyiladi bema'ni mutatsiya. Agar bunday to'xtash signali gen ichida (uning axborot qismida) hosil bo'lsa, u holda bu joyda protein sintezi paytida jarayon doimiy ravishda to'xtatiladi - oqsilning faqat birinchi (to'xtash signalidan oldin) qismi sintezlanadi. Ushbu patologiyaga ega bo'lgan odam oqsil etishmasligini boshdan kechiradi va bu etishmovchilik bilan bog'liq alomatlar paydo bo'ladi. Masalan, bunday mutatsiya gemoglobin beta zanjirini kodlovchi genda aniqlangan. Qisqartirilgan faol bo'lmagan gemoglobin zanjiri sintezlanadi, u tezda yo'q qilinadi. Natijada, beta zanjiridan mahrum bo'lgan gemoglobin molekulasi hosil bo'ladi. Bunday molekulaning o'z vazifalarini to'liq bajarishi dargumon. Gemolitik anemiya shaklida rivojlanadigan jiddiy kasallik (beta-nol talassemiya, yunoncha "Talas" so'zidan - O'rta er dengizi, bu kasallik birinchi marta kashf etilgan).
To'xtash kodonlarining ta'sir qilish mexanizmi hissiy kodonlarning ta'sir qilish mexanizmidan farq qiladi. Bu aminokislotalarni kodlaydigan barcha kodonlar uchun mos keladigan tRNKlar topilganligidan kelib chiqadi. Bema'ni kodonlar uchun tRNK topilmadi. Binobarin, tRNK oqsil sintezini to'xtatish jarayonida ishtirok etmaydi.
KodonAVG (ba'zan bakteriyalarda GUG) nafaqat metionin va valin aminokislotalarini kodlaydi, balkieshittirish tashabbuskori .
b. Degeneratsiya yoki ortiqchalik.
64 ta uchlikdan 61 tasi 20 ta aminokislotalarni kodlaydi. Aminokislotalar sonidan uch baravar ko'pligi ma'lumot uzatishda ikkita kodlash variantidan foydalanish mumkinligini ko'rsatadi. Birinchidan, 20 ta aminokislotalarni kodlashda 64 ta kodonning hammasi ham ishtirok eta olmaydi, faqat 20 ta, ikkinchidan, aminokislotalarni bir nechta kodonlar bilan kodlash mumkin. Tadqiqotlar shuni ko'rsatdiki, tabiat oxirgi variantdan foydalangan.
Uning afzalligi aniq. Agar 64 ta variantdan atigi 20 tasi aminokislotalarni kodlashda ishtirok etgan bo'lsa, u holda 44 ta uchlik (64 tadan) kodlanmagan bo'lib qoladi, ya'ni. ma'nosiz (bema'ni kodonlar). Ilgari biz mutatsiya natijasida kodlovchi tripletni bema'ni kodonga aylantirish hujayra hayoti uchun qanchalik xavfli ekanligini ta'kidlagan edik - bu sezilarli darajada buzadi. normal ish RNK polimerazalari, oxir-oqibat kasalliklarning rivojlanishiga olib keladi. Hozirgi vaqtda bizning genomimizdagi uchta kodon bema'nilikdir, ammo endi ma'nosiz kodonlar soni taxminan 15 baravar ko'paysa nima bo'lishini tasavvur qiling. Bunday vaziyatda oddiy kodonlarning ma'nosiz kodonlarga o'tishi beqiyos darajada yuqori bo'lishi aniq.
Bitta aminokislota bir nechta uchlik bilan kodlangan kod degenerativ yoki ortiqcha deb ataladi. Deyarli har bir aminokislota bir nechta kodonlarga ega. Shunday qilib, aminokislota leysin oltita uchlik - UUA, UUG, TSUU, TsUC, TsUA, TsUG bilan kodlanishi mumkin. Valin to'rtta uchlik, fenilalanin ikkita va faqat kodlangan triptofan va metionin bitta kodon bilan kodlangan. Turli xil belgilar bilan bir xil ma'lumotlarni yozib olish bilan bog'liq xususiyat deyiladi degeneratsiya.
Bitta aminokislota uchun belgilangan kodonlar soni oqsillarda aminokislotalarning paydo bo'lish chastotasi bilan yaxshi bog'liq.
Va bu, ehtimol, tasodifiy emas. Proteinda aminokislotalarning paydo bo'lish chastotasi qanchalik yuqori bo'lsa, genomda ushbu aminokislotalarning kodoni qanchalik tez-tez namoyon bo'lsa, mutagen omillar ta'sirida uning shikastlanish ehtimoli shunchalik yuqori bo'ladi. Shu sababli, mutatsiyaga uchragan kodon, agar u juda degeneratsiyalangan bo'lsa, bir xil aminokislotalarni kodlash imkoniyati ko'proq ekanligi aniq. Shu nuqtai nazardan qaraganda, genetik kodning degeneratsiyasi inson genomini shikastlanishdan himoya qiluvchi mexanizmdir.
Shuni ta'kidlash kerakki, degeneratsiya atamasi molekulyar genetikada boshqa ma'noda qo'llaniladi. Shunday qilib, kodondagi ma'lumotlarning asosiy qismi birinchi ikkita nukleotidda joylashgan bo'lib, kodonning uchinchi pozitsiyasidagi asos unchalik ahamiyatga ega emas. Ushbu hodisa "uchinchi bazaning degeneratsiyasi" deb ataladi. Oxirgi xususiyat mutatsiyalarning ta'sirini kamaytiradi. Masalan, qizil qon hujayralarining asosiy vazifasi kislorodni o'pkadan to'qimalarga va karbonat angidridni to'qimalardan o'pkaga o'tkazish ekanligi ma'lum. Bu funktsiyani nafas olish pigmenti - gemoglobin bajaradi, u eritrotsitning butun sitoplazmasini to'ldiradi. U tegishli gen tomonidan kodlangan oqsil qismi - globindan iborat. Proteindan tashqari, gemoglobin molekulasi tarkibida temir o'z ichiga olgan gem mavjud. Globin genlaridagi mutatsiyalar paydo bo'lishiga olib keladi turli xil variantlar gemoglobinlar. Ko'pincha mutatsiyalar bilan bog'liq bir nukleotidni boshqasi bilan almashtirish va genda yangi kodon paydo bo'lishi, bu gemoglobin polipeptid zanjirida yangi aminokislotalarni kodlashi mumkin. Tripletda mutatsiya natijasida har qanday nukleotid almashtirilishi mumkin - birinchi, ikkinchi yoki uchinchi. Globin genlarining yaxlitligiga ta'sir qiluvchi bir necha yuz mutatsiyalar ma'lum. Yaqin 400 ulardan gendagi yagona nukleotidlarning almashinishi va polipeptiddagi tegishli aminokislotalarning almashinuvi bilan bog'liq. Faqat shulardan 100 almashtirish gemoglobinning beqarorligiga va engildan o'ta og'irgacha bo'lgan turli xil kasalliklarga olib keladi. 300 (taxminan 64%) almashtirish mutatsiyasi gemoglobin funktsiyasiga ta'sir qilmaydi va patologiyaga olib kelmaydi. Buning sabablaridan biri serin, leysin, prolin, arginin va boshqa ba'zi aminokislotalarni kodlovchi tripletda uchinchi nukleotidning almashinishi sinonimik kodonning paydo bo'lishiga olib keladigan yuqorida aytib o'tilgan "uchinchi asosning degeneratsiyasi" dir. bir xil aminokislotalarni kodlash. Bunday mutatsiya fenotipik tarzda o'zini namoyon qilmaydi. Aksincha, 100% hollarda birinchi yoki ikkinchi nukleotidning tripletdagi har qanday almashinuvi yangi gemoglobin variantining paydo bo'lishiga olib keladi. Ammo bu holatda ham jiddiy fenotipik buzilishlar bo'lmasligi mumkin. Buning sababi gemoglobindagi aminokislotani birinchisiga o'xshash boshqasi bilan almashtirishdir. fizik va kimyoviy xossalari. Misol uchun, agar gidrofil xossaga ega bo'lgan aminokislota boshqa aminokislota bilan almashtirilsa, lekin bir xil xususiyatlarga ega.
Gemoglobin gemning temir porfirin guruhidan (kislorod va karbonat angidrid molekulalari biriktirilgan) va oqsil - globindan iborat. Katta yoshli gemoglobin (HbA) ikkita bir xil gemoglobinni o'z ichiga oladi - zanjirlar va ikkita -zanjirlar. Molekula -zanjirda 141 ta aminokislota qoldiqlari mavjud; -zanjir - 146, - Va -zanjirlar ko'p aminokislotalar qoldiqlarida farqlanadi. Har bir globin zanjirining aminokislotalar ketma-ketligi o'z geni tomonidan kodlangan. Genni kodlash -zanjir 16-xromosomaning qisqa qo'lida joylashgan; -gen - 11-xromosomaning qisqa qo'lida. Genlarni kodlashda almashtirish -birinchi yoki ikkinchi nukleotidning gemoglobin zanjiri deyarli har doim oqsilda yangi aminokislotalarning paydo bo'lishiga, gemoglobin funktsiyalarining buzilishiga va bemor uchun jiddiy oqibatlarga olib keladi. Masalan, CAU (histidin) uchliklaridan birida "C" ni "Y" bilan almashtirish boshqa aminokislota - tirozinni kodlaydigan yangi uchlik UAU paydo bo'lishiga olib keladi 63-pozitsiyada xuddi shunday almashtirish -gistidin polipeptidining tirozinga zanjiri gemoglobinning beqarorlashishiga olib keladi. Kasallik methemoglobinemiya rivojlanadi. Mutatsiya natijasida glutamik kislotani 6-pog'onada valin bilan almashtirish -zanjir eng og'ir kasallikning sababi - o'roqsimon hujayrali anemiya. Keling, qayg'uli ro'yxatni davom ettirmaylik. Shuni ta'kidlash kerakki, dastlabki ikkita nukleotidni almashtirganda, avvalgisiga fizik-kimyoviy xususiyatlarga o'xshash aminokislota paydo bo'lishi mumkin. Shunday qilib, glutamik kislotani (GAA) kodlovchi tripletlardan birida 2-nukleotidni almashtirish. -"U" bilan zanjir valinni kodlaydigan yangi triplet (GUA) paydo bo'lishiga olib keladi va birinchi nukleotidni "A" bilan almashtirish lizin aminokislotasini kodlaydigan AAA tripletini hosil qiladi. Glutamik kislota va lizin fizik-kimyoviy xossalari bo'yicha o'xshash - ikkalasi ham gidrofildir. Valin hidrofobik aminokislotadir. Shuning uchun gidrofil glutamik kislotani hidrofobik valin bilan almashtirish gemoglobinning xususiyatlarini sezilarli darajada o'zgartiradi, bu oxir-oqibat o'roqsimon hujayrali anemiya rivojlanishiga olib keladi, gidrofil glutamik kislotani gidrofil lizin bilan almashtirish esa gemoglobin funktsiyasini kamroq darajada o'zgartiradi - bemorlarda engil shakl rivojlanadi. anemiyadan. Uchinchi asosni almashtirish natijasida yangi triplet avvalgisi kabi bir xil aminokislotalarni kodlashi mumkin. Misol uchun, agar CAC tripletida urasil sitozin bilan almashtirilgan bo'lsa va CAC triplet paydo bo'lsa, u holda odamlarda deyarli hech qanday fenotipik o'zgarishlar aniqlanmaydi. Bu tushunarli, chunki Ikkala triplet ham bir xil aminokislota - histidinni kodlaydi.
Xulosa qilib shuni ta'kidlash joizki, genetik kodning degeneratsiyasi va umumiy biologik nuqtai nazardan uchinchi bazaning degeneratsiyasi. himoya mexanizmlari, ular DNK va RNKning noyob tuzilishida evolyutsiyaga xosdir.
V. Aniqlik.
Har bir triplet (bema'nilikdan tashqari) faqat bitta aminokislotani kodlaydi. Shunday qilib, kodon - aminokislota yo'nalishida genetik kod bir ma'noli, aminokislota - kodon yo'nalishi bo'yicha u noaniq (degeneratsiya).
Aniq
Aminokislota kodoni
Degeneratsiya
Va bu holda, genetik kodda noaniqlik zarurati aniq. Boshqa variantda, bir xil kodonni tarjima qilganda, oqsil zanjiriga turli xil aminokislotalar kiritiladi va natijada, birlamchi tuzilmalari va funktsiyalari turlicha bo'lgan oqsillar hosil bo'ladi. Hujayra metabolizmi "bir gen - bir nechta polipeptidlar" ish rejimiga o'tadi. Bunday vaziyatda genlarning tartibga solish funktsiyasi butunlay yo'qolishi aniq.
g
DNK va mRNK dan ma'lumotni o'qish faqat bitta yo'nalishda sodir bo'ladi. Polarite bor muhim yuqori tartibli tuzilmalarni (ikkilamchi, uchinchi darajali va boshqalarni) aniqlash. Ilgari biz tuzilmalarni aytdik pastki tartib yuqori darajadagi tuzilmalarni aniqlash. Sintezlangan RNK zanjiri DNK molekulasini yoki polipeptid zanjiri ribosomani tark etishi bilanoq oqsillardagi uchinchi darajali tuzilma va yuqori tartibli tuzilmalar hosil bo'ladi. RNK yoki polipeptidning erkin uchi uchinchi darajali tuzilishga ega bo'lsa, zanjirning ikkinchi uchi DNKda (agar RNK transkripsiyalangan bo'lsa) yoki ribosomada (agar polipeptid transkripsiyalangan bo'lsa) sintezlanishda davom etadi.
Shuning uchun ma'lumotni o'qishning bir yo'nalishli jarayoni (RNK va oqsil sintezi paytida) nafaqat sintez qilingan moddadagi nukleotidlar yoki aminokislotalarning ketma-ketligini aniqlash uchun, balki ikkilamchi, uchinchi darajali va boshqalarni qat'iy aniqlash uchun muhimdir. tuzilmalar.
d. Bir-biriga mos kelmaslik.
Kod bir-biriga o'xshash yoki bir-biriga mos kelmasligi mumkin. Aksariyat organizmlar bir-birining ustiga chiqmaydigan kodga ega. Ba'zi faglarda bir-biriga o'xshash kod mavjud.
Bir-biriga yopishmaydigan kodning mohiyati shundan iboratki, bir kodonning nukleotidi bir vaqtning o'zida boshqa kodonning nukleotidi bo'la olmaydi. Agar kod bir-biriga o'xshash bo'lsa, etti nukleotidlar ketma-ketligi (GCUGCUG) bir-birining ustiga chiqmaydigan koddagi kabi ikkita aminokislotani (alanin-alanin) (33-rasm, A) emas, balki uchtasini (agar mavjud bo'lsa) kodlashi mumkin edi. umumiy bitta nukleotid) (33-rasm, B) yoki beshta (agar ikkita nukleotid umumiy bo'lsa) (33-rasmga qarang, C). So'nggi ikki holatda, har qanday nukleotidning mutatsiyasi ikki, uch va hokazo ketma-ketlikning buzilishiga olib keladi. aminokislotalar.
Biroq, bitta nukleotidning mutatsiyasi har doim bitta aminokislotaning polipeptid tarkibiga kirishini buzishi aniqlangan. Bu kodning bir-biriga mos kelmasligini ko'rsatadigan muhim dalil.
Buni 34-rasmda tushuntiramiz. Qalin chiziqlar bir-birining ustiga chiqmagan va bir-birining ustiga chiquvchi kod bo'lgan taqdirda aminokislotalarni kodlovchi uchliklarni ko'rsatadi. Tajribalar genetik kodning bir-biriga mos kelmasligini aniq ko'rsatdi. Tajriba tafsilotlariga kirmasdan, shuni ta'kidlaymizki, agar siz uchinchi nukleotidni nukleotidlar ketma-ketligida almashtirsangiz (34-rasmga qarang).U (yulduzcha bilan belgilangan) boshqa narsaga:
1. Bir-biriga mos kelmaydigan kod bilan ushbu ketma-ketlik bilan boshqariladigan oqsil bitta (birinchi) aminokislota o'rnini bosadi (yulduzcha bilan belgilangan).
2. A variantida bir-biriga o'xshash kod bo'lsa, ikkita (birinchi va ikkinchi) aminokislotalarda (yulduzcha bilan belgilangan) almashtirish sodir bo'ladi. B variantiga ko'ra, almashtirish uchta aminokislotaga ta'sir qiladi (yulduzcha bilan belgilangan).
Biroq, ko'plab tajribalar shuni ko'rsatdiki, DNKdagi bitta nukleotid buzilganda, oqsildagi buzilish har doim bir-biriga mos kelmaydigan kod uchun xos bo'lgan faqat bitta aminokislotaga ta'sir qiladi.
GZUGZUG GZUGZUG GZUGZUG
GCU GCU GCU UGC GCU GCU GCU UGC GCU GCU GCU
*** *** *** *** *** ***
Alanin - Alanin Ala - Cis - Ley Ala - Ley - Ley - Ala - Ley
A B C
Bir-biriga mos kelmaydigan kod Bir-biriga mos keladigan kod
Guruch. 34. Genomda bir-biriga mos kelmaydigan kod mavjudligini tushuntiruvchi diagramma (matndagi tushuntirish).
Genetik kodning bir-biriga mos kelmasligi boshqa xususiyat bilan bog'liq - ma'lumotni o'qish ma'lum bir nuqtadan boshlanadi - boshlash signali. mRNKdagi bunday boshlash signali AUG metioninni kodlovchi kodondir.
Shuni ta'kidlash kerakki, odamlarda hali ham oz sonli genlar mavjud umumiy qoida va bir-biriga yopishadi.
e. Kompaktlik.
Kodonlar orasida tinish belgilari yo'q. Boshqacha qilib aytganda, tripletlar bir-biridan, masalan, bitta ma'nosiz nukleotid bilan ajratilmaydi. Genetik kodda "tinish belgilari" yo'qligi tajribalarda isbotlangan.
va. Ko'p qirralilik.
Kod Yerda yashovchi barcha organizmlar uchun bir xil. Genetik kodning universalligi to'g'ridan-to'g'ri dalil DNK ketma-ketligini mos keladigan oqsil ketma-ketliklari bilan taqqoslash orqali olingan. Ma'lum bo'lishicha, barcha bakterial va eukaryotik genomlar bir xil kod qiymatlari to'plamidan foydalanadi. Istisnolar bor, lekin ko'p emas.
Genetik kodning universalligiga birinchi istisnolar ba'zi hayvonlar turlarining mitoxondriyalarida topilgan. Bu triptofan aminokislotasini kodlovchi UGG kodon bilan bir xil o'qiydigan UGA terminator kodoniga tegishli edi. Umumjahonlikdan boshqa kam uchraydigan og'ishlar ham topildi.
DNK kod tizimi.
DNK genetik kodi 64 ta nukleotid tripletidan iborat. Bu tripletlar kodonlar deb ataladi. Har bir kodon oqsil sintezida ishlatiladigan 20 ta aminokislotadan birini kodlaydi. Bu kodda biroz ortiqchalikni beradi: ko'pchilik aminokislotalar bir nechta kodon tomonidan kodlangan.
Bitta kodon o'zaro bog'liq bo'lgan ikkita funktsiyani bajaradi: u tarjimaning boshlanishi haqida signal beradi va o'sib borayotgan polipeptid zanjiriga metionin (Met) aminokislotalarining kiritilishini kodlaydi. DNK kodlash tizimi shunday yaratilganki, genetik kod RNK kodonlari yoki DNK kodonlari sifatida ifodalanishi mumkin. RNK kodonlari RNKda (mRNK) topiladi va bu kodonlar polipeptidlar sintezi paytida ma'lumotni o'qiy oladi (bu jarayon translatsiya deb ataladi). Ammo har bir mRNK molekulasi tegishli gendan transkripsiyada nukleotidlar ketma-ketligini oladi.
Ikki aminokislotadan tashqari (Met va Trp) hammasi 2 dan 6 gacha turli kodonlar bilan kodlanishi mumkin. Biroq, ko'pchilik organizmlarning genomi shuni ko'rsatadiki, ba'zi kodonlar boshqalarga nisbatan afzalroqdir. Odamlarda, masalan, alanin GCC tomonidan GCGga qaraganda to'rt marta ko'proq kodlanadi. Bu, ehtimol, ba'zi kodonlar uchun tarjima apparatining (masalan, ribosoma) yuqori tarjima samaradorligini ko'rsatadi.

Genetik kod deyarli universaldir. Xuddi shu kodonlar aminokislotalarning bir xil bo'limiga biriktirilgan va bir xil boshlash va to'xtash signallari hayvonlar, o'simliklar va mikroorganizmlarda deyarli bir xil. Biroq, ba'zi istisnolar topildi. Ko'pchilik uchta to'xtash kodonidan birini yoki ikkitasini aminokislotalarga tayinlashni o'z ichiga oladi.
Gen- muayyan xususiyat yoki xususiyatning rivojlanishini nazorat qiluvchi irsiyatning tarkibiy va funktsional birligi. Ko'payish jarayonida ota-onalar o'z avlodlariga genlar to'plamini o'tkazadilar: Simashkevich E.A., Gavrilova Yu.A., Bogomazova O.V (2011).
Hozirgi vaqtda molekulyar biologiyada genlar DNKning bo'limlari ekanligi aniqlandi, ular qandaydir integral ma'lumotni - bitta protein molekulasi yoki bitta RNK molekulasining tuzilishi haqida. Bu va boshqa funktsional molekulalar tananing rivojlanishi, o'sishi va faoliyatini belgilaydi.
Shu bilan birga, har bir gen gen ekspressiyasini tartibga solishda bevosita ishtirok etuvchi promotorlar kabi bir qator o'ziga xos tartibga soluvchi DNK ketma-ketliklari bilan tavsiflanadi. Tartibga soluvchi ketma-ketliklar oqsilni kodlaydigan ochiq o'qish ramkasiga yoki promotorlarda bo'lgani kabi RNK ketma-ketligining boshlanishiga yaqin joyda joylashgan bo'lishi mumkin (deb ataladi). cis cis-tartibga solish elementlari) va ko'p million tayanch juftlari (nukleotidlar) masofalarida, kuchaytirgichlar, izolyatorlar va bostiruvchilar (ba'zan shunday tasniflanadi) misolida bo'lgani kabi trans-tartibga solish elementlari, ingliz tili. trans-regulyatsiya elementlari). Shunday qilib, gen tushunchasi faqat DNKning kodlash mintaqasi bilan chegaralanib qolmaydi, balki tartibga solish ketma-ketliklarini ham o'z ichiga olgan kengroq tushunchadir.
Dastlab atama gen diskret irsiy axborotni uzatish uchun nazariy birlik sifatida paydo bo'ldi. Biologiya tarixi qaysi molekulalar irsiy ma'lumotni tashuvchisi bo'lishi mumkinligi haqidagi bahslarni eslaydi. Ko'pgina tadqiqotchilar faqat oqsillar bunday tashuvchi bo'lishi mumkinligiga ishonishgan, chunki ularning tuzilishi (20 ta aminokislotalar) yaratishga imkon beradi. ko'proq imkoniyatlar faqat tashkil topgan DNK tuzilishiga qaraganda to'rt xil nukleotidlar. Keyinchalik bu DNKni o'z ichiga olganligi eksperimental ravishda isbotlangan irsiy ma'lumotlar, bu molekulyar biologiyaning markaziy dogmasi sifatida ifodalangan.
Genlar mutatsiyaga uchrashi mumkin - DNK zanjiridagi nukleotidlar ketma-ketligidagi tasodifiy yoki maqsadli o'zgarishlar. Mutatsiyalar ketma-ketlikning o'zgarishiga va shuning uchun o'zgarishga olib kelishi mumkin biologik xususiyatlar oqsil yoki RNK, bu o'z navbatida tananing umumiy yoki mahalliy o'zgarishi yoki anormal ishlashiga olib kelishi mumkin. Bunday mutatsiyalar ba'zi hollarda patogen hisoblanadi, chunki ular kasallik yoki embrion darajasida o'limga olib keladi. Biroq, nukleotidlar ketma-ketligidagi barcha o'zgarishlar oqsil strukturasining o'zgarishiga (genetik kodning degeneratsiyasi ta'siri tufayli) yoki ketma-ketlikning sezilarli o'zgarishiga olib kelmaydi va patogen emas. Xususan, inson genomi bir nukleotid polimorfizmi va nusxa sonining o'zgarishi bilan tavsiflanadi. raqam o'zgarishlarini nusxalash), masalan, o'chirish va dublikatsiyalar, bu butun inson nukleotidlari ketma-ketligining taxminan 1% ni tashkil qiladi. Yagona nukleotid polimorfizmlari, xususan, bitta genning turli allellarini belgilaydi.
Har bir DNK zanjirini tashkil etuvchi monomerlar murakkabdir organik birikmalar, shu jumladan azotli asoslar: adenin (A) yoki timin (T) yoki sitozin (C) yoki guanin (G), pentaatomik shakar-pentoza-deoksiriboz, undan keyin DNKning o'zi, shuningdek, fosforik kislota qoldig'i bu birikmalar nukleotidlar deb ataladi.
Gen xususiyatlari
- diskretlik - genlarning aralashmasligi;
- barqarorlik - strukturani saqlab qolish qobiliyati;
- labillik - qayta-qayta mutatsiya qilish qobiliyati;
- ko'p allelizm - ko'plab genlar populyatsiyada bir nechta molekulyar shakllarda mavjud;
- allellik - diploid organizmlar genotipida genning faqat ikkita shakli mavjud;
- o'ziga xoslik - har bir gen o'ziga xos xususiyatni kodlaydi;
- pleiotropiya - genning ko'p ta'siri;
- ekspressivlik - belgidagi genning ifodalanish darajasi;
- penetratsiya - fenotipdagi genning namoyon bo'lish chastotasi;
- amplifikatsiya - gen nusxalari sonini ko'paytirish.
Tasniflash
- Strukturaviy genlar genomning o'ziga xos tarkibiy qismlari bo'lib, ma'lum bir protein yoki ma'lum turdagi RNKni kodlaydigan yagona ketma-ketlikni ifodalaydi. (Shuningdek, genlar maqolasiga qarang uy xo'jaligi).
- Funktsional genlar - strukturaviy genlarning faoliyatini tartibga soladi.
Genetik kod- nukleotidlar ketma-ketligi yordamida oqsillarning aminokislotalar ketma-ketligini kodlashning barcha tirik organizmlarga xos bo'lgan usuli.
DNK to'rtta nukleotiddan foydalanadi - adenin (A), guanin (G), sitozin (C), timin (T), rus adabiyotida A, G, C va T harflari bilan belgilanadi. Bu harflar alifboning alifbosini tashkil qiladi. genetik kod. RNK bir xil nukleotidlardan foydalanadi, timin bundan mustasno, u xuddi shunday nukleotid - urasil bilan almashtiriladi, u U harfi bilan belgilanadi (rus adabiyotida U). DNK va RNK molekulalarida nukleotidlar zanjir bo'lib joylashadi va shu tariqa genetik harflar ketma-ketligi olinadi.
Genetik kod
Tabiatda oqsillarni qurish uchun 20 xil aminokislotalar ishlatiladi. Har bir oqsil qat'iy belgilangan ketma-ketlikda aminokislotalarning zanjiri yoki bir nechta zanjiridir. Bu ketma-ketlik oqsilning tuzilishini va shuning uchun uning hammasini belgilaydi biologik xossalari. Aminokislotalar to'plami ham deyarli barcha tirik organizmlar uchun universaldir.
Tirik hujayralarda genetik ma'lumotni amalga oshirish (ya'ni gen tomonidan kodlangan oqsil sintezi) ikkita matritsa jarayoni yordamida amalga oshiriladi: transkripsiya (ya'ni DNK matritsasida mRNK sintezi) va genetik kodni tarjima qilish. aminokislotalar ketma-ketligiga (mRNKda polipeptid zanjirining sintezi). 20 ta aminokislotalarni kodlash uchun ketma-ket uchta nukleotid, shuningdek, oqsil ketma-ketligining tugashini ko'rsatadigan to'xtash signali etarli. Uch nukleotiddan iborat to'plamga triplet deyiladi. Qabul qilingan qisqartmalar, aminokislotalar va kodonlarga mos keladigan, rasmda ko'rsatilgan.
Xususiyatlari
- Uchlik- kodning mazmunli birligi uchta nukleotidning (uchlik yoki kodon) birikmasidir.
- Davomiylik- uchlik o'rtasida tinish belgilari yo'q, ya'ni ma'lumotlar uzluksiz o'qiladi.
- Bir-biriga mos kelmaslik- bir xil nukleotid bir vaqtning o'zida ikki yoki undan ortiq tripletlarning bir qismi bo'lishi mumkin emas (bir nechta ramka almashinuvi oqsillarini kodlaydigan viruslar, mitoxondriyalar va bakteriyalarning bir-biriga o'xshash ba'zi genlari uchun kuzatilmaydi).
- O'ziga xoslik (o'ziga xoslik)- ma'lum bir kodon faqat bitta aminokislotaga to'g'ri keladi (ammo UGA kodonida mavjud Euplotes crassus ikkita aminokislotalarni kodlaydi - sistein va selenosistein)
- Degeneratsiya (ortiqchalik)- bir aminokislotaga bir nechta kodon mos kelishi mumkin.
- Ko'p qirralilik- genetik kod turli darajadagi murakkablikdagi organizmlarda bir xil ishlaydi - viruslardan tortib to odamlargacha (gen muhandislik usullari bunga asoslanadi; "Standart genetik kodning o'zgarishlari" bo'limidagi jadvalda ko'rsatilgan bir qator istisnolar mavjud; quyida).
- Shovqinga qarshi immunitet- kodlangan aminokislotalar sinfining o'zgarishiga olib kelmaydigan nukleotid almashtirish mutatsiyalari deyiladi. konservativ; kodlangan aminokislota sinfining o'zgarishiga olib keladigan nukleotidlarni almashtirish mutatsiyalari deyiladi radikal.
Oqsil biosintezi va uning bosqichlari
Protein biosintezi- mRNK va tRNK molekulalari ishtirokida tirik organizmlar hujayralari ribosomalarida yuzaga keladigan aminokislotalar qoldiqlaridan polipeptid zanjirini sintez qilishning murakkab ko'p bosqichli jarayoni.
Protein biosintezini transkripsiya, qayta ishlash va translatsiya bosqichlariga bo'lish mumkin. Transkripsiya paytida DNK molekulalarida shifrlangan genetik ma'lumot o'qiladi va bu ma'lumot mRNK molekulalariga yoziladi. Bir qator ketma-ket ishlov berish bosqichlarida, keyingi bosqichlarda keraksiz bo'lgan ba'zi fragmentlar mRNKdan chiqariladi va nukleotidlar ketma-ketligi tahrirlanadi. Kod yadrodan ribosomalarga o'tkazilgandan so'ng, oqsil molekulalarining haqiqiy sintezi o'sib borayotgan polipeptid zanjiriga individual aminokislotalar qoldiqlarini biriktirish orqali sodir bo'ladi.
Transkripsiya va translatsiya o'rtasida mRNK molekulasi polipeptid zanjiri sintezi uchun ishlaydigan matritsaning etukligini ta'minlaydigan bir qator ketma-ket o'zgarishlarga uchraydi. 5n-uchiga qalpoq, 3n-uchiga esa poli-A dumi biriktirilgan, bu esa mRNKning ishlash muddatini oshiradi. Eukaryotik hujayrada qayta ishlashning paydo bo'lishi bilan DNK nukleotidlarining bitta ketma-ketligi bilan kodlangan ko'proq turli xil oqsillarni olish uchun gen eksonlarini birlashtirish mumkin bo'ldi - alternativ birlashma.
Tarjima messenjer RNKda kodlangan ma'lumotlarga muvofiq polipeptid zanjirining sintezidan iborat. Aminokislotalar ketma-ketligi yordamida tartibga solinadi transport Aminokislotalar bilan komplekslar hosil qiluvchi RNK (tRNK) - aminoatsil-tRNK. Har bir aminokislota o'z tRNKsiga ega, u mRNK kodoniga "mos keladigan" tegishli antikodonga ega. Translyatsiya paytida ribosoma mRNK bo'ylab harakatlanadi va buning natijasida polipeptid zanjiri o'sadi. Protein biosintezi uchun energiya ATP tomonidan ta'minlanadi.
Keyin tayyor oqsil molekulasi ribosomadan ajraladi va unga ko'chiriladi To'g'ri joy hujayralar. O'zlarining faol holatiga erishish uchun ba'zi oqsillar qo'shimcha post-translational modifikatsiyani talab qiladi.