Первичная структура белка пептидная связь. Первичная, вторичная, третичная и четвертичная структуры белка. Дополнительные структуры белка
Объединение аминокислот через пептидные связи создает линейную полипептидную цепь, которая называется первичной структурой белка
Учитывая, что в синтезе белков принимает участие 20 аминокислот и средний белок содержит 500 аминокислотных остатков, то можно говорить о невообразимом количестве потенциально возможных белков. В организме человека обнаружено около 100 тысяч различных белков.
К примеру, 2 аминокислоты (аланин и серин) образуют 2 пептида Ала-Сер и Сер-Ала; 3 аминокислоты дадут уже 6 вариантов трипептида; 20 аминокислот – 1018 различных пептидов длиной всего 20 аминокислот (при условии, что каждая аминокислота используется только один раз).
Самый большой из известных в настоящее время белков - титин - является компонентом саркомеров миоцита, молекулярная масса его различных изоформ находится в интервале от 3000 до 3700 кДа. Титин камбаловидной мышцы человека состоит из 38138 аминокислот.
Первичная структура белков, т.е. последовательность аминокислот в нем, программируется последовательностью нуклеотидов в ДНК. Выпадение, вставка, замена нуклеотида в ДНК приводит к изменению аминокислотного состава и, следовательно, структуры синтезируемого белка.
Участок белковой цепи длиной в 6 аминокислот (Сер-Цис-Тир-Лей-Глу-Ала)
(пептидные связи выделены желтым фоном, аминокислоты - рамкой)
Если изменение последовательности аминокислот носит не летальный характер, а приспособительный или хотя бы нейтральный, то новый белок может передаться по наследству и остаться в популяции. В результате возникают новые белки с похожими функциями. Такое явление называется полиморфизм белков.
Для многих белков обнаруживается ярко выраженный консерватизм структуры. Например, гормон инсулин у человека отличается от бычьего только на три аминокислоты, от свиного – на одну аминокислоту (аланин вместо треонина).
Последовательность и соотношение аминокислот в первичной структуре определяет формирование вторичной , третичной и четвертичной структур.
Генотипическая гетерогенность
В результате того, что каждый ген у человека имеется в двух копиях (аллелях) и может подвергаться мутациям (замена, делеция, вставка) и рекомбинациям, серьезно не затрагивающим функцию кодируемого белка, то возникает полиморфизм генов и, соответственно, полиморфизм белков. Возникают целые семейства родственных белков, обладающих схожими, но неодинаковыми свойствами и функцией.
Например, существует около 300 разных типов гемоглобина , часть из них является необходимой на разных этапах онтогенеза: например, HbP – эмбриональный, образуется в первый месяц развития, HbF – фетальный, необходим на более поздних сроках развития плода, HbA и HbA2 – гемоглобин взрослых. Разнообразие обеспечивается полиморфизмом глобиновых цепей: в гемоглобине P присутствуют 2ξ и 2ε цепи, в HbF – 2α- и 2γ- цепи, в HbА – 2α- и 2β-цепи, в HbА2 – 2α- и 2δ-цепи.
При серповидноклеточной анемии в шестом положении β-цепи гемоглобина происходит замена глутаминовой кислоты на валин. Это приводит к синтезу гемоглобина S (HbS) – такого гемоглобина, который в дезоксиформе полимеризуется и образует тяжи. В результате эритроциты деформируются, приобретают форму серпа (банана), теряют эластичность и при прохождении через капилляры разрушаются. Это в итоге приводит к снижению оксигенации тканей и их некрозу.
Группы крови АВ0
зависят от строения особого углевода на мембране эритроцитов. Различия в строении углевода обусловлены разной специфичностью и активностью фермента гликозил-трансферазы
, способного модифицировать исходный олигосахарид. Фермент имеет три варианта и осуществляет присоединение к олигосахариду мембран эритроцитов либо N-ацетилгалактозы, либо галактозы, либо фермент не присоединяет дополнительные сахаридные группы (группа 0).
В результате лица с группой крови А0 на эритроците имеют олигосахарид с присоединенным к нему N-ацетилгалактозамином, с группой крови В0 – олигосахарид с галактозой, 00 – имеют только "чистый" олигосахарид, с группой крови АВ – олигосахарид и с N-ацетилгалактозамином, и с галактозой.
Как известно, белки - основа зарождения жизни на нашей планете. По именно коацерватная капля, состоящая из молекул пептидов, стала основой зарождения живого. Это и не вызывает сомнений, ведь анализ внутреннего состава любого представителя биомассы показывает, что эти вещества есть во всем: растениях, животных, микроорганизмах, грибах, вирусах. Причем они очень разнообразны и макромолекулярны по природе.
Названий у этих структур четыре, все они являются синонимами:
- белки;
- протеины;
- полипептиды;
- пептиды.
Белковые молекулы
Их количество поистине неисчислимо. При этом все белковые молекулы можно разделить на две большие группы:
- простые - состоят только из аминокислотных последовательностей, соединенных пептидными связями;
- сложные - строение и структура белка характеризуются дополнительными протолитическими (простетическими) группами, называемыми еще кофакторами.
При этом сложные молекулы также имеют свою классификацию.
Градация сложных пептидов
- Гликопротеиды - тесно связанные соединения белка и углевода. В структуру молекулы вплетаются простетические группы мукополисахаридов.
- Липопротеиды - комплексное соединение из белка и липида.
- Металлопротеиды - в качестве простетической группы выступают ионы металлов (железо, марганец, медь и другие).
- Нуклеопротеиды - связь белка и нуклеиновых кислот (ДНК, РНК).
- Фосфопротеиды - конформация протеина и остатка ортофосфорной кислоты.
- Хромопротеиды - очень схожи с металлопротеидами, однако элемент, входящий в состав простетической группы, представляет собой целый окрашенный комплекс (красный - гемоглобин, зеленый - хлорофилл и так далее).
У каждой рассмотренной группы строение и свойства белков различны. Функции, которые они выполняют, также варьируются в зависимости от типа молекулы.

Химическое строение белков
С данной точки зрения протеины - это длинная, массивная цепь аминокислотных остатков, соединяющихся между собой специфическими связями, называемыми пептидными. От боковых структур кислот отходят ответвления - радикалы. Такое строение молекулы было открыто Э. Фишером в начале XXI века.
Позже более подробно были изучены белки, строение и функции белков. Стало ясно, что аминокислот, образующих структуру пептида, всего 20, но они способны комбинироваться самым разным способом. Отсюда и разнообразие полипептидных структур. Кроме того, в процессе жизнедеятельности и выполнения своих функций белки способны претерпевать ряд химических превращений. В результате они меняют структуру, и появляется уже совсем новый тип соединения.
Чтобы разорвать пептидную связь, то есть нарушить белок, строение цепей, нужно подобрать очень жесткие условия (действие высоких температур, кислот или щелочей, катализатора). Это объясняется высокой прочностью в молекуле, а именно в пептидной группе.

Обнаружение белковой структуры в условиях лаборатории проводится при помощи биуретовой реакции - воздействия на полипептид свежеосажденным (II). Комплекс пептидной группы и иона меди дает ярко-фиолетовую окраску.
Существует четыре основные структурные организации, каждая из которых имеет свои особенности строения белков.
Уровни организации: первичная структура
Как уже упоминалось выше, пептид - это последовательность аминокислотных остатков с включениями, коферментами или же без них. Так вот первичной называют такую структуру молекулы, которая является природной, естественной, представляет собой истинно аминокислоты, соединенные пептидными связями, и больше ничего. То есть полипептид линейного строения. При этом особенности строения белков такого плана - в том, что такое сочетание кислот является определяющим для выполнения функций белковой молекулы. Благодаря наличию данных особенностей возможно не только идентифицировать пептид, но и предсказать свойства и роль совершенно нового, еще не открытого. Примеры пептидов, обладающих природным первичным строением, - инсулин, пепсин, химотрипсин и другие.

Вторичная конформация
Строение и свойства белков этой категории несколько меняются. Такая структура может сформироваться изначально от природы либо при воздействии на первичную жестким гидролизом, температурой или иными условиями.
Данная конформация имеет три разновидности:
- Ровные, правильные, стереорегулярные витки, построенные из остатков аминокислот, которые закручиваются вокруг основной оси соединения. Удерживаются вместе только возникающими между кислородом одной пептидной группировки и водородом другой. Причем строение считается правильным из-за того, что витки равномерно повторяются через каждые 4 звена. Такая структура может быть как левозакрученной, так и правозакрученной. Но в большинстве известных белков преобладает правовращающий изомер. Такие конформации принято называть альфа-структурами.
- Состав и строение белков следующего типа отличается от предыдущего тем, что водородные связи образуются не между рядом стоящими по одной стороне молекулы остатками, а между значительно удаленными, причем на достаточно большое расстояние. По этой причине вся структура принимает вид нескольких волнообразных, извитых змейкой полипептидных цепочек. Есть одна особенность, которую должен проявлять белок. Строение аминокислот на ответвлениях должно быть максимально коротким, как у глицина или аланина, например. Этот тип вторичной конформации носит название бета-листов за способность будто слипаться при образовании общей структуры.
- Относящееся к третьему типу строение белка биология обозначает как сложные, разноразбросанные, неупорядоченные фрагменты, не обладающие стереорегулярностью и способные изменять структуру под воздействием внешних условий.
Примеров белков, имеющих вторичную структуру от природы, не выявлено.

Третичное образование
Это достаточно сложная конформация, имеющая название "глобула". Что собой представляет такой белок? Строение его основывается на вторичной структуре, однако добавляются новые типы взаимодействий между атомами группировок, и вся молекула словно сворачивается, ориентируясь, таким образом, на то, чтобы гидрофильные группировки были направлены внутрь глобулы, а гидрофобные - наружу.
Этим объясняется заряд белковой молекулы в коллоидных растворах воды. Какие же типы взаимодействий здесь присутствуют?
- Водородные связи - остаются без изменений между теми же самыми частями, что и во вторичной структуре.
- взаимодействия - возникают при растворении полипептида в воде.
- Ионные притяжения - образуются между разнозаряженными группами аминокислотных остатков (радикалов).
- Ковалентные взаимодействия - способны формироваться между конкретными кислотными участками - молекулами цистеина, вернее, их хвостами.
Таким образом, состав и строение белков, обладающих третичной структурой, можно описать как свернутые в глобулы полипептидные цепи, удерживающие и стабилизирующие свою конформацию за счет разных типов химических взаимодействий. Примеры таких пептидов: фосфоглицераткеназа, тРНК, альфа-кератин, фиброин шелка и другие.
Четвертичная структура
Это одна из самых сложных глобул, которую образуют белки. Строение и функции белков подобного плана очень многогранны и специфичны.
Что собой представляет такая конформация? Это несколько (в некоторых случаях десятки) крупных и мелких полипептидных цепей, которые формируются независимо друг от друга. Но затем за счет тех же взаимодействий, что мы рассматривали для третичной структуры, все эти пептиды скручиваются и переплетаются между собой. Таким образом получаются сложные конформационные глобулы, которые могут содержать и атомы металлов, и липидные группировки, и углеводные. Примеры таких белков: ДНК-полимераза, белковая оболочка табачного вируса, гемоглобин и другие.

Все рассмотренные нами структуры пептидов имеют свои методы идентификации в лабораторных условиях, основанные на современных возможностях использования хроматографии, центрифугирования, электронной и оптической микроскопии и высоких компьютерных технологиях.
Выполняемые функции
Строение и функции белков тесно коррелируют друг с другом. То есть каждый пептид играет определенную роль, уникальную и специфическую. Встречаются и такие, которые способны выполнять в одной живой клетке сразу несколько значительных операций. Однако можно в обобщенном виде выразить основные функции белковых молекул в организмах живых существ:
- Обеспечение движения. Одноклеточные организмы, либо органеллы, или некоторые виды клеток способны к передвижениям, сокращениям, перемещениям. Это обеспечивается белками, входящими в состав структуры их двигательного аппарата: ресничек, жгутиков, цитоплазматической мембраны. Если же говорить о неспособных к перемещениям клетках, то белки могут способствовать их сокращению (миозин мышц).
- Питательная или резервная функция. Представляет собой накопление белковых молекул в яйцеклетках, зародышах и семенах растений для дальнейшего восполнения недостающих питательных веществ. При расщеплении пептиды дают аминокислоты и биологически активные вещества, которые необходимы для нормального развития живых организмов.
- Энергетическая функция. Помимо углеводов, силы организму могут давать и белки. При распаде 1 г пептида высвобождается 17,6 кДж полезной энергии в форме аденозинтрифосфорной кислоты (АТФ), которая расходуется на процессы жизнедеятельности.
- Сигнальная и Заключается в осуществлении тщательного контроля за происходящими процессами и передачи сигналов от клеток к тканям, от них к органам, от последних к системам и так далее. Типичным примером может служить инсулин, который строго фиксирует количество глюкозы в крови.
- Рецепторная функция. Осуществляется путем изменения конформации пептида с одной стороны мембраны и вовлечения в реструктуризацию другого конца. При этом и происходит передача сигнала и необходимой информации. Чаще всего такие белки встраиваются в цитоплазматические мембраны клеток и осуществляют строгий контроль над всеми веществами, проходящими через нее. Также оповещают о химических и физических изменениях окружающей среды.
- Транспортная функция пептидов. Ее осуществляют белки-каналы и белки-переносчики. Роль их очевидна - транспортировка необходимых молекул к местам с низкой концентрацией из частей с высокой. Типичным примером служит перенос кислорода и диоксида углерода по органам и тканям белком гемоглобином. Ими же осуществляется доставка соединений с невысокой молекулярной массой через мембрану клетки внутрь.
- Структурная функция. Одна из важнейших из тех, которые выполняет белок. Строение всех клеток, их органелл обеспечивается именно пептидами. Они подобно каркасу задают форму и структуру. Кроме того, они же ее поддерживают и видоизменяют в случае необходимости. Поэтому для роста и развития всем живым организмам необходимы белки в рационе питания. К таким пептидам можно отнести эластин, тубулин, коллаген, актин, кератин и другие.
- Каталитическая функция. Ее выполняют ферменты. Многочисленные и разнообразные, они ускоряют все химические и биохимические реакции в организме. Без их участия обычное яблоко в желудке смогло бы перевариться только за два дня, с большой вероятностью загнив при этом. Под действием каталазы, пероксидазы и других ферментов этот процесс происходит за два часа. В целом именно благодаря такой роли белков осуществляется анаболизм и катаболизм, то есть пластический и

Защитная роль
Существует несколько типов угроз, от которых белки призваны оберегать организм.
Во-первых, травмирующих реагентов, газов, молекул, веществ различного спектра действия. Пептиды способны вступать с ними в химическое взаимодействие, переводя в безобидную форму или же просто нейтрализуя.
Во-вторых, физическая угроза со стороны ран - если белок фибриноген вовремя не трансформируется в фибрин на месте травмы, то кровь не свернется, а значит, закупорка не произойдет. Затем, наоборот, понадобится пептид плазмин, способный сгусток рассосать и восстановить проходимость сосуда.
В-третьих, угроза иммунитету. Строение и значение белков, формирующих иммунную защиту, крайне важны. Антитела, иммуноглобулины, интерфероны - все это важные и значимые элементы лимфатической и иммунной системы человека. Любая чужеродная частица, вредоносная молекула, отмершая часть клетки или целая структура подвергается немедленному исследованию со стороны пептидного соединения. Именно поэтому человек может самостоятельно, без помощи лекарственных средств, ежедневно защищать себя от инфекций и несложных вирусов.
Физические свойства
Строение белка клетки весьма специфично и зависит от выполняемой функции. А вот физические свойства всех пептидов схожи и сводятся к следующим характеристикам.
- Вес молекулы - до 1000000 Дальтон.
- В водном растворе формируют коллоидные системы. Там структура приобретает заряд, способный варьироваться в зависимости от кислотности среды.
- При воздействии жестких условий (облучение, кислота или щелочь, температура и так далее) способны переходить на другие уровни конформаций, то есть денатурировать. Данный процесс в 90% случаев необратим. Однако существует и обратный сдвиг - ренатурация.
Это основные свойства физической характеристики пептидов.
П ЕРВИЧНАЯ СТРУКТУРА БЕЛКОВ
Первичная структура белка несет информацию о его пространственной структуре.
1.Аминокислотные остатки в пептидной цепи белков чередуются не случайным образом, а распо-ложены в определенном порядке. Линейная после-довательность аминокислотных остатков в полипеп-тидной цепи называется первичной структурой белка.
2. Первичная структура каждого индивидуально-го белка закодирована в молекуле ДНК (участке, называемом геном) и реализуется в ходе транс-крипции (переписывания информации на мРНК) и трансляции (синтез пептидной цепи).
3. Каждый из 50 000 индивидуальных белков ор-ганизма человека имеет уникальную для данного индивидуального белка первичную структуру. Все молекулы индивидуального белка (например, аль-бумина) имеют одинаковое чередование амино-кислотных остатков, отличающее альбумин от лю-бого другого индивидуального белка.
4. Последовательность аминокислотных остат-ков в пептидной цепи можно рассматривать как
форму запи
си некоторой информации.
Эта информация диктует пространственную ук-ладку длинной линейной пептидной цепи в более компактную трехмерную структуру.
КОНФОРМАЦИЯ БЕЛКОВ
1. Линейные полипептидные цепи индивидуаль-ных белков за счет взаимодействия функциональ-ных групп аминокислот приобретают определен-ную пространственную трехмерную структуру, или конформацию. В глобулярных белках различают
два основных типа конформации
пептидных цепей: вторичную и третичную структуры.
ВТОРИЧНАЯ СТРУКТУРА БЕЛКОВ
2. Вторичная структура белков - это пространст-венная структура, образующаяся в результате взаимодействий между функциональными груп- пами пептидного остова. При этом пептидная цепь может приобретать регулярные структуры двух типов: ос-спирали и р-структуры.
Рис. 1.2. Вторичная структура белка — а-спираль.
В ос-спирали водородные связи образуются между атомом кислорода карбоксильной группы и водородом амидного азота пептидного остова через 4 аминокислоты; боковые цепи аминокислотных остатков располагаются по периферии спирали, не участвуя в образовании водородных связей, фор-мирующих вторичную структуру (рис. 1.2).
Большие объемные остатки или остатки с одина-ковыми отталкивающимися зарядами препятству- ют формированию а-спирали.
Остаток пролина прерывает а-спираль благодаря его кольцевой структуре и невозможности образо-вания водородной связи из-за отсутствия водорода у атома азота в пептидной цепи.
B -Структура формируется между линейными областями одной полипептидной цепи, образуя при этом складки, или между разными полипеп-тидными цепями. Полипептидные цепи или их части могут формировать параллельные (N- и С-концы взаимодействующих пептидных цепей совпадают) или антипараллельные (N- и С-концы взаимодействующих пептидных цепей лежат в противоположных направлениях) р-структуры (рис. 1.3).
В белках также встречаются области с нерегу-лярной вторичной структурой, которые называ-ются беспорядочными клубками, хотя эти структу-ры не так сильно изменяются от одной молекулы белка к другой.
ТРЕТИЧНАЯ СТРУКТУРА БЕЛКОВ
3. Третичная структура белка — это трехмерная пространственная структура, образующаяся за счет взаимодействий между радикалами аминокислот, которые могут располагаться на значительном рас-стоянии друг от друга в пептидной цепи.
Рис. 1.3. Антипараллельная (бета-структура.)
.jpg)
Гидрофобные радикалы аминокислот имеют тенденцию к объединению внутри глобулярной структуры белков с помощью так называемых гид- рофобных взаимодействий и межмолекулярных ван-дер-ваальсовых сил, образуя плотное гидро-фобное ядро. Гидрофильные ионизированные и неионизированные радикалы аминокислот в ос-новном расположены на поверхности белка и оп-ределяют его растворимость в воде.
Гидрофильные аминокислоты, оказавшиеся внут-ри гидрофобного ядра, могут взаимодействовать друг с другом с помощью ионных и водородных свя-зей (рис. 1.4).
.jpg)
Рис. 1.4. Типы связей, возникающие между радикалами аминокислот при формировании третичной структуры белка. 1 — ионная связь; 2 — водородная связь; 3 — гидрофобные взаимодействия; 4 — дисульфидная связь.
|
|
.jpg)
Рис. 1.5. Дисульфидные связи в структуре инсулина человека.
Ионные, водородные и гидрофобные связи отно-сятся к числу слабых: их энергия ненамного пре-вышает энергию теплового движения молекул при комнатной температуре.
Конформация белка поддерживается за счет воз-никновения множества таких слабых связей.
Конформационная лабильность белков — это спо-собность белков к небольшим изменениям кон-формации за счет разрыва одних и образования других слабых связей.
Третичная структура некоторых белков стабили-зирована дисульфидными связями, образующимися за счет взаимодействия SH-групп двух остатков цистеина.
Большинство внутриклеточных белков не имеет ковалентных дисульфидных связей. Их наличие характерно для секретируемых клеткой белков, на-пример дисульфидные связи имеются в молекулах инсулина, иммуноглобулинов.
Инсулин — белковый гормон, синтезирующийся в р-клетках поджелудочной железы. Секретируется клетками в ответ на повышение концентрации глю-козы в крови. В структуре инсулина имеются 2 ди-сульфидные связи, соединяющие 2 полипептидные А- и В-цепи, и 1 дисульфидная связь внутри А-цепи (рис. 1.5).
Особенности вторичной структуры белков ока-зывают влияние на характер межрадикальных вза-имодействий и третичную структуру.
4. Некоторый специфический порядок чередова-ния вторичных структур наблюдается во многих разных по структуре и функциям белках и носит название супервторичной структуры.
Такие упорядоченные структуры часто обозначают как структурные мотивы, которые имеют специфические названия: «а-спираль—поворот—а-спи-раль», «лейциновая застежка-молния», «цинковые пальцы», «структура Р-бочонка» и др.
По наличию а-спиралей и р-структур глобуляр-ные белки могут быть разделены на 4 категории:
1.В первую категорию включены белки, в кото-рых имеются только а-спирали, например миогло-бин и гемоглобин (рис. 1.6).
2. Во вторую категорию включены белки, в кото-рых имеются а-спирали и (3-структуры. При этом а- и (3-структуры часто образуют однотипные со-четания, встречающиеся в разных индивидуаль-ных белках.
Пример. Супервторичная структура типа Р-бочонка.

Фермент триозофосфатизомераза имеет супер-вторичную структуру типа Р-бочонка, где каждая (3-структура расположена внутри р-бочонка и свя-зана с а-спиральным участком полипептидной цепи, находящимся на поверхности молекулы (рис. 1.7, а).
Рис. 1.7. Супервторичная структура типа р-бочонка.
а — триозофосфатизомераза; б — домен пиру ватки назы.
.jpg)
Такая же супервторичная структура обнаружена в одном из доменов молекулы фермента пируваткиназы (рис. 1.7, б). Доменом называют часть молеку-лы, по структуре напоминающую самостоятель-ный глобулярный белок.
Еще один пример формирования супервторич-ной структуры, имеющей Р-структуры и ос-спира-ли. В одном из доменов лактатдегидрогеназы (ЛДГ) и фосфоглицераткиназы в центре располо-жены Р-структуры полипептидной цепи в виде скрученного листа и каждая р-структура связана с а-спиральным участком, расположенным на по-верхности молекулы (рис. 1.8).
Рис. 1.8. Вторичная структура, характерная для многих фер- ментов.
а -домен лактатдегидрогеназы; б— домен фосфоглицераткиназы.
.jpg) 3. В третью категорию включены белки, имею-
щие только вторичную р-структуру. Такие структу-ры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе (рис. 1.9).
3. В третью категорию включены белки, имею-
щие только вторичную р-структуру. Такие структу-ры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе (рис. 1.9).
Рис. 1.9. Вторичная структура константного домена им-муноглобулина (а)
и фермента супероксиддисмутазы (б).
.jpg)
4. В четвертую категорию включены белки, имеющие в своем составе лишь незначительное ко-личество регулярных вторичных структур. К таким белкам можно отнести небольшие богатые цисти-ном белки или металлопротеины.
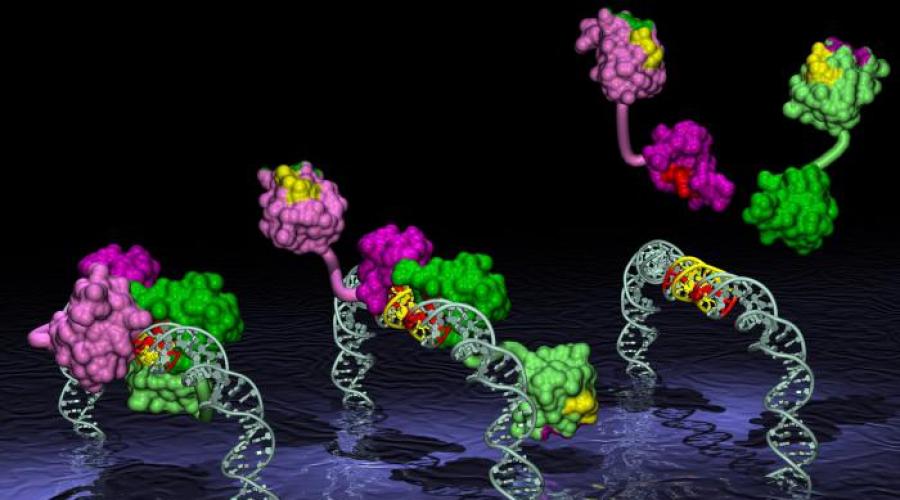
В ДНК-связывающих белках имеются общие виды супервторичных структур: «ос-спираль—поворот— ос-спираль», «лейциновая застежка-молния», «цинко- вые пальцы». ДНК-связывающие белки содержат центр связывания, комплементарный участку ДНК с определенной нуклеотидной последовательностью. Эти белки участвуют в регуляции действия генов.
«а-
Спираль—поворот—а-спираль»
Рис. 1.10. Связывание супервторичной
структуры «а-спи-раль—поворот—а-спираль»
в большой бороздке Д
|
|
.jpg)
.jpg) шая бороздка хорошо
при-способлена для связывания белков, имеющих не-большие ос-спиральные участки.
шая бороздка хорошо
при-способлена для связывания белков, имеющих не-большие ос-спиральные участки.
В данный структурный мотив входят 2 ос-спирали: одна более короткая, другая более длинная, соеди-ненные поворотом полипептидной цепи (рис. 1.10).
Более короткая а-спираль располагается попе-рек бороздки ДНК, а более длинная а-спираль на-ходится в большой бороздке, образуя нековалент-ные специфические связи радикалов аминокислот с нуклеотидами ДНК.
Часто белки, имеющие такую структуру, образу-ют димеры, в результате олигомерный белок имеет 2 супервторичные структуры.
Они располагаются на определенном расстоянии друг от друга и выступают над поверхностью белка (рис. 1.11).Две такие структуры могут связываться с ДНК в смежных областях больших бороздок
без значи-тельных изменений в структуре белков.
«Цинковый палец»
«Цинковый палец» — фрагмент белка, содержа-щий около 20 аминокислотных остатков (рис. 1.12).
Атом цинка связан с радикалами 4 аминокислот: 2 остатков цистеина и 2 — гистидина.
В некоторых случаях вместо остатков гистидина находятся остатки цистеина.
Рис. 1.12. Структура участка ДНК-связывающихбелков в форме «цинкового пальца».
.jpg)
Этот участок белка образует а-спираль, которая может специфично связываться с регуляторными участками большой бороздки ДНК.
Специфичность связывания индивидуального регуляторного ДНК-связывающего белка зависит от последовательности аминокислотных остатков, расположенных в области «цинкового пальца».
«Лейциновая застежка-молния»
Взаимодействующие белки имеют а-спиральный участок, содержащий по крайней мере 4 ос-татка лейцина.
Лейциновые остатки расположены через 6 ами-нокислот один от другого.
Так как каждый виток а-спирали содержит 3,6-аминокислотного остатка, радикалы лейцина находятся на поверхности каждого второго витка.
Лейциновые остатки а-спирали одного белка могут взаимодействовать с лейциновыми остатка-ми другого белка (гидрофобные взаимодействия), соединяя их вместе (рис. 1.13).
.jpg) Многие ДНК-связывающие белки взаимодейст-вуют с ДНК в виде олигомерных структур, где субъединицы связываются друг с другом «лейци-новыми застежками». Примером таких белков мо-гут служить гистоны.
Многие ДНК-связывающие белки взаимодейст-вуют с ДНК в виде олигомерных структур, где субъединицы связываются друг с другом «лейци-новыми застежками». Примером таких белков мо-гут служить гистоны.
Гистоны — ядерные белки, в состав которых вхо-дит большое количество положительно заряжен-ных аминокислот — аргинина и лизина (до 80%).
Молекулы гистонов объединяются в олигомер-ные комплексы, содержащие 8 мономеров с по-мощью «лейциновых застежек», несмотря на силь-ный положительный заряд этих молекул.
Резюме. Все молекулы индивидуального белка, имеющие идентичную первичную структуру, при-обретают в растворе одинаковую конформацию.
Таким образом, характер пространственной уклад-ки пептидной цепи определяется аминокислотным составом и чередованием аминокислотных остатков в цепи. Следовательно, конформация — такая же специфическая характеристика индивидуального белка, как и первичная структура.
03. Строение белковой молекулы (первичная, вторичная, третичная, четвертичная). Типы связей. Связь структуры с функцией
Каждый белок характеризуется специфической аминокислотной последовательностью и индивидуальной пространственной структурой (конформацией). На долю белков приходится не менее 50% сухой массы органических соединений животной клетки. В организме человека насчитывается до 5 млн. различных видов белков. Белковая молекула может состоять из одной или нескольких цепей, содержащих от пятидесяти до нескольких сотен аминокислотных остатков. Молекулы, содержащие менее пятидесяти остатков, относят к пептидам . В состав многих молекул входят остатки цистеина, дисульфидные связи которых ковалентно связывают участки одной или нескольких цепей. В нативном состоянии белковые макромолекулы обладают специфической конформацией. Характерная для данного белка конформация определяется:
- последовательностью аминокислотных остатков и стабилизируется водородными связями между пептидными и боковыми группами аминокислотных остатков,
- электростатическими и гидрофобными взаимодействиями.
Первичная структура белка. Пептидная связь образуется при реакции аминогруппы одной аминокислоты и карбоксильной группы другой с выделением молекулы воды :
CH3-CH(NH2)-COOH + CH3- CH(NH2)-COOH ^ CH3-CH(NH2)-CO- NH-(CH3) СН-COOH + H2O
Связанные пептидной связью аминокислоты образуют полипептидную цепь . Пептидная связь имеет плоскостную структуру:
- атомы С, О и N находятся в sp -гибридизации;
- у атома N имеется р-орбиталь с неподеленной парой электронов;
- образуется р-п-сопряженная система, приводящая к укорочению связи С-N (0,132 нм) и ограничению вращения (барьер вращения составляет ~63 кДж/моль).
Пептидная связь имеет преимущественно трансконфигурацию относительно плоскости пептидной связи. Подобное строение пептидной связи сказывается на формировании вторичной и третичной структуры белка. Пептидная связь жесткая, ковалентная, генетически детерминированная. В структурных формулах изображается в виде одинарной связи, однако на самом деле эта связь между углеродом и азотом носит характер частично двойной связи. Это вызвано различной электроотрицательностью атомов С, N и O. Вокруг пептидной связи вращение невозможно, все четыре атома лежат в одной плоскости, т.е. компланарны. Вращение же других связей вокруг полипептидного остова достаточно свободно. Последовательность аминокислот для каждого белка уникальна и закреплена генетически.
По числу аминокислотных остатков, входящих в молекулы пептидов, различают дипептиды, трипептиды, тетрапептиды и т.д. Пептиды, содержащие до десяти аминокислотных остатков, называются олигопептидами , содержащие более десяти аминокислотных остатков - полипептидами . Природные полипептиды с молекулярной массой более 6000 называются белками.
Вторичная структура - это пространственное расположение полипептидной цепочки в виде а-спирали или Р-складчатости безотносительно к типам боковых радикалов и их конформации. Л. Полинг и Р. Кори предложили модель вторичной структуры белка в виде а-спирали , в которой водородные связи замыкаются между каждой первой и четвертой аминокислотой, что позволяет сохранять нативную структуру белка, осуществлять простейшие функции, защищать от разрушения. В образовании водородных связей принимают участие все пептидные группы, что обеспечивает максимальную стабильность, снижает гидрофильность и увеличивает гидрофобность белковой молекулы. а-спираль образуется самопроизвольно и является наиболее устойчивой конформацией, отвечающей минимуму свободной энергии. Наиболее распространенным элементом вторичной структуры является правая а-спираль (aR ). Пептидная цепь здесь изгибается винтообразно. Ha каждый виток приходится 3,6 аминокислотного остатка, шаг винта, т.е. минимальное расстояние между двумя эквивалентными точками, составляет 0,54 нм; а-спираль стабилизирована почти линейными водородными связями между NH-группой и СО-группой четвертого по счету аминокислотного остатка. Неполярные или амфифильные а-спирали с 5-6 витками часто обеспечивают заякоривание белков в биологических мембранах (трансмембранные спирали). B складчатых структурах также образуются поперечные межцепочечные водородные связи Если цепи ориентированы в противоположных направлениях, структура называется антипараллельным складчатым листом (ва); если цепи ориентированы в одном направлении, структура называется параллельным складчатым листом (вп). Кроме регулярных в полипептидных цепях есть еще и нерегулярные вторичные структуры, т.е. стандартные структуры, не образующие длинных периодических систем. Это - в-изгибы они называются так потому, что часто стягивают верхушки соседних в-тяжей в антипараллельных в-шпильках). В изгибы обычно входит около половины остатков, не опавших в регулярные структуры белков.
Связи, стабилизирующие третичную структуру :
- электростатические силы притяжения между R-группами, несущими противоположно заряженные ионогенные группы (ионные связи);
- водородные связи между полярными (гидрофильными) R-группами;
- гидрофобные взаимодействия между неполярными (гидрофобными) R-группами;
- дисульфидные связи между радикалами двух молекул цистеина.
Эти связи ковалентные. Они повышают стабильность третичной структуры, но не всегда являются обязательными для правильного скручивания молекулы. В ряде белков они могут вообще отсутствовать.
Третичная структура - уникальное для каждого белка расположение в пространстве полипептидной цепи, зависящее от количества и чередования аминокислот, т.е. предопределенное первичной структурой белка. Конфигурация белковых молекул может быть фибриллярной и глобулярной . Третичная структура многих белков составляется из нескольких компактных глобул, называемых доменами . Между собой домены обычно бывают связаны тонкими перемычками вытянутыми аморфными полипептидными цепями Кроме того, в белках встречаются мотивы укладки полипептидной цепи, похожие на орнаменты на индейских и греческих вазах: мотив меандра, мотив греческого ключа, мотив зигзага-"молнии" При свертывании белковой глобулы значительная часть (не менее половины) гидрофобных радикалов аминокислотных остатков оказывается скрытой от контакта с окружающей белок водой. Происходит образование своеобразных внутримолекулярных «гидрофобных ядер ». В них особенно представлены объемные остатки лейцина, изолейцина, фенилаланина, валина. С появлением третичной структуры у белка появляются новые свойства - биологические . В частности, проявление каталитических свойств связано с наличием у белка третичной структуры. Фибриллярные белки - белки, имеющие вытянутую нитевидную структуру. Большинство фибриллярных белков не растворяется в воде, имеют большую молекулярную массу и высокорегулярную пространственную структуру, которая стабилизируется, главным образом, взаимодействиями (в том числе и ковалентными) между различными полипептидными цепями. Полипептидные цепи многих фибриллярных белков расположены параллельно друг другу вдоль одной оси и образуют длинные волокна (фибриллы) или слои . Глобулярные белки - белки, в молекулах которых полипептидные цепи плотно свёрнуты в компактные шарообразные структуры - глобулы (третичные структуры белка).
Четвертичная структура - это надмолекулярное образование, состоящее из двух и более полипептидных цепей, связанных между собой нековалентно, а водородными связями, электростатическими, дипольдипольные и гидрофобными взаимодействиями между остатками аминокислот, находящихся на поверхности. Каждый из белков-участников третичной структуры при образовании четвертичной структуры называют субъединицей или протомером . Образовавшуюся молекулу называют олигомером, или мультимером . Олигомерные белки чаще построены из четного количества протомеров с одинаковыми или разными молекулярными массами. В образовании четвертичной структуры белка принимают участие те же связи, что и при образовании третичной структуры, за исключением ковалентных. Характерной особенностью белков с четвертичной структурой является их способность к самосборке. Взаимодействие протомеров осуществляется с высокой специфичностью, благодаря образованию десятка слабых связей между контактными поверхностями субъединиц, поэтому ошибки при формировании четвертичной структуры белков исключены.
Практически все белки-ферменты имеют четвертичную структуру и состоят, как правило, из четного числа протомеров (двух, четырех, шести, восьми). Четвертичная структура белка подразумевает такое объединение белков третичной структуры, при котором появляются новые биологические свойства, не характерные для белка в третичной структуре.
Пространственная конфигурация белка т.е. третичная и четвертичная структуры называется конформацией . Если полипептидную цепь взять за концы, растянуть ее и затем отпустить, то она всякий раз будет свертываться в одну и ту же структуру, характерную для этого вида полипептида. В то же время из сказанного, очевидно, следует, что, изменив всего лишь одну аминокислоту в каком-либо полипептиде, мы получим молекулу с совершенно иной структурой, а значит и с иными свойствами.
По химическому составу все белки делят на простые , состоящие только из аминокислотных остатков, и сложные . Сложные могут включать ионы металла (металлопротеины) или пигмент (хромопротеины), образовывать прочные комплексы с липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеины), а также ковалентно связывать остаток фосфорной кислоты (фосфопротеины), углевода (гликопротеины).
Простые белки подразделяют на:
- фибриллярные, растворимые в воде (актин, миозин) и нерастворимые (кератин, эластин, коллаген),
- глобулярные (альбумины, глобулины, протамины, гистоны, проламины).
Как показало изучение свойств белков в растворах, макромолекулы белков имеют форму компактных шаров (глобул) или вытянутых структур – фибрилл. Исследования показали, что в укладке пептидной цепи нет ничего случайного или хаотичного. Она свертывается упорядоченно, для каждого белка определенным образом. Полярные боковые группы аминокислот стремятся расположиться на поверхности глобулы, где могут взаимодействовать с водой, а неполярные группы располагаются внутри.
Для того, чтобы разобраться в замысловатой укладке (архитектонике) белковой макромолекулы, следует рассмотреть в ней несколько уровней организации .
Первичной структурой белка называется полная последовательность аминокислотных остатков в полипептидной цепи.
Она определяется генотипом, т.е. генами организма. В первичной структуре все связи между аминокислотными остатками являются ковалентными и, следовательно, прочными. Разные белки отличаются друг от друга по первичной структуре: кератин имеет одну последовательность аминокислот, пепсин – другую, соматотропин (гормон роста) – третью и т.д. В первичной структуре белка можно различить N-конец цепочки, содержащий свободную NH 2 -группу, и С-конец, содержащий свободную СООН-группу.
Первым белком, у которого была выявлена аминокислотная последовательность, стал гормон инсулин. Исследования проводились в Кембриджском университете Ф.Сэнгером в 1944–1954 гг. Было выявлено, что молекула инсулина состоит из двух полипептидных цепей (из 21 и 30 аминокислотных остатков), удерживаемых друг около друга дисульфидными мостиками.
Однако молекула белка в виде цепи аминокислот, последовательно соединенных пептидными связями, еще не способна выполнять специфические функции. Для этого необходим более высокий уровень структурной организации, выражающийся в усложнении пространственного расположения мономеров.
Вторичная структура белка представлена спиралью, в которую закручивается полипептидная цепь. Группы N–H и С=О, входящие в пептидную связь, заметно поляризованы: азот обладает большей электроотрицательностью, чем водород, а кислород – большей, чем углерод.
Кислород группы С=О может образовывать водородные связи с водородом группы N–H (разумеется, расположенной в другой пептидной связи).
Одной из разновидностей вторичной структуры является -спираль , где каждый атом кислорода связан с атомом водорода четвертой по ходу спирали NH-группы.
Любопытно, что эта сложная красивая структура сперва была предсказана известным биохимиком Л.Полингом теоретически и лишь потом обнаружена экспериментально.
Альтернативная вторичная структура -слой (или складчатый слой) имеет водородную связь между звеньями соседних полипептидных цепей.
Третичная структура белка
представляет собой сложную трехмерную пространственную упаковку
-спиралей и -слоев. Эта трехмерная структура устанавливается за счет взаимодействия радикалов аминокислот, между которыми могут возникнуть связи несколько типов:
1) ионные , возникающие за счет электростатического взаимодействия между отрицательно и положительно заряженными боковыми группами;
2) гидрофобные («не любящие воду»), устанавливающиеся за счет стремления неполярных радикалов объединяться друг с другом, а не смешиваться с окружающей их водной средой;
3) дисульфидные , которые образуются между атомами серы SH-групп двух остатков аминокислоты цистеина. Эти S–S связи по своей природе являются ковалентными;
4) водородные , которые также возникают за счет взаимодействия между атомами радикалов.
Третичная структура белка не является конечной. Для некоторых белков, чаще всего регуляторных, характерна четвертичная структура, необходимая им для эффективного выполнения функции.
Четвертичная структура представлена ассоциантом, состоящим из нескольких полипептидных цепей. Например, сложная молекула гемоглобина состоит из двух -субъединиц (141 аминокислотный остаток) и двух -субъединиц (146 аминокислотных остатков). Каждая субъединица связана с молекулой железосодержащего гема. В результате их объединения образуется функционирующая молекула гемоглобина. Только в такой упаковке гемоглобин работает полноценно, то есть способен переносить кислород. Четвертичная структура стабилизируется теми же связями, что и третичная.
Пространственная конфигурация белка т.е. третичная и четвертичная структуры называется конформацией . Конформация белка определяется его первичной структурой: белковая цепочка с определенной последовательностью аминокислот самопроизвольно укладывается с образованием природной пространственной конфигурации. Это получило название «самосборка белковой молекулы». Если полипептидную цепь взять за концы, растянуть ее и затем отпустить, то она всякий раз будет свертываться в одну и ту же структуру, характерную для этого вида полипептида.
В то же время из сказанного, очевидно, следует, что, изменив всего лишь одну аминокислоту в каком-либо полипептиде, мы получим молекулу с совершенно иной структурой, а значит и с иными свойствами.