L'accepteur de co2 en c4 de la voie photosynthétique est. Caractéristiques comparatives des plantes -C3 et -C4. Lipides : classification et leur rôle dans la plante
Voies alternatives de photosynthèse.
C 3 voies s'est avéré être non seulement dans la transformation du Co 2 mais dans la transformation en phase sombre. Les changements dans les conditions climatiques extérieures et la composition atmosphérique dans différentes parties de la Terre ont provoqué l'émergence de continents au climat chaud et sec (déserts), ce qui ne pouvait qu'affecter le monde végétal. La spécificité des espèces est sans aucun doute le reflet du caractère unique du type physiologique de chaque espèce végétale. Depuis les années 50, on s'intéresse aux processus de photosynthèse, de respiration, de transpiration et de transport de substances. Il s'est avéré que les conditions environnementales déterminent les types structurels et biochimiques de la photosynthèse végétale.
Au début des années 60 du XXe siècle, on a constaté que certaines plantes poussant sous les tropiques ou en émergeant, comme le maïs, la canne à sucre, le sorgho, le mil, représentants de la famille des chénopodes du semi-désert et désert, au total 18 familles. Ces plantes se trouvent dans diverses familles et genres, mais il n’existe pas une seule famille ou un seul genre composé uniquement de plantes C4. Les premiers produits de l'assimilation du CO 2 comportent ici 4 atomes de carbone, c'est pourquoi ces plantes sont appelées « plantes C4 ». Dans ces plantes, le produit primaire stable du CO 2 assimilé n'est pas le PHA, mais l'acide malique, et dans un certain nombre de plantes, l'acide aspartique, contenant non pas 3, mais 4 atomes de carbone, dans lequel le marqueur radioactif 14 CO 2 a été découvert (Yu .S Karpilov, 1960, 1969 ; M.D. Hatch et S.R. Ainsi, une voie essentiellement nouvelle d'assimilation du CO 2 a été découverte, appelée la « voie C 4 » de la photosynthèse.
Ces plantes ont une structure foliaire anatomique spécifique. Ils possèdent deux types de cellules photosynthétiques : les cellules de la gaine, qui recouvrent radialement les faisceaux vasculaires de la feuille, et les cellules mésophylles du tissu colonnaire et spongieux. Cela peut servir de signe diagnostique pour déterminer les plantes C4. Il existe également des différences dans la structure de ces cellules : dans les cellules de la gaine des chloroplastes, il n'y a pas de grana, elles sont agranales, et dans les cellules mésophylles, tous les chloroplastes sont granal. Ces deux types de cellules ne sont pas physiologiquement équivalents et se spécialisent dans des fonctions différentes dans la conversion du CO 2 absorbé.
Les cellules de la gaine remplissent la fonction principale ; le cycle de Calvin s'y déroule, de la même manière qu'il se produit dans les plantes à C 3 par la photosynthèse, c'est-à-dire l'ensemble du processus de conversion du dioxyde de carbone à travers toutes les étapes du cycle de Calvin. Les cellules du mésophylle des feuilles jouent un rôle de soutien, en pompant le CO 2 pour le cycle de Calvin. Ainsi, chez les plantes dotées de la voie C 4. Il y a deux stades, se déroulant dans différentes cellules de la feuille. Pour un chloroplaste agranal d'une cellule de la gaine, il existe 10 cellules mésophylles granales, dans lesquelles se produit une voie de photophosphorylation non cyclique. Cela garantit une augmentation de la photosynthèse même dans des conditions de températures très élevées et de stomates presque fermés, ce qui a également un effet positif sur le régime hydrique des plantes, puisque la consommation d'eau est réduite. Les deux processus sont spatialement séparés, c'est-à-dire que l'assimilation du CO 2 se produit dans les cellules du mésophylle, et la formation de sucres et la régénération de l'accepteur dans les cellules pariétales. Mais ces deux processus se produisent simultanément, dans la lumière.
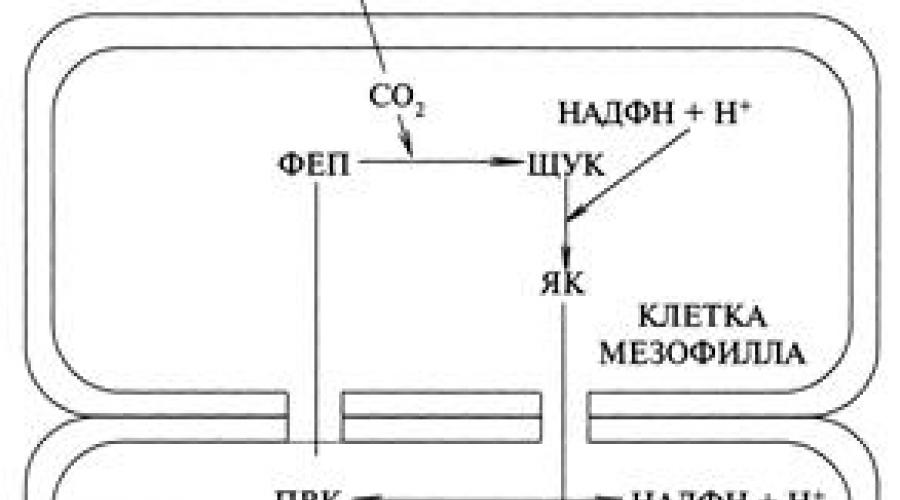
La première réaction du cycle C 4 - la voie de la photosynthèse se produit dans le mésophylle de la feuille, dans laquelle l'acide phosphoénol-pyruvique (PEP) est l'accepteur de dioxyde de carbone. Dans ces cellules, la PEP carboxylase est très active, liant intensément le CO 2 en un composé organique (Fig.)

Cycle photosynthétique C 4 – voies de la photosynthèse
PEP - acide phosphoénolpyruvique ;
PCHUK - acide oxaloacétique ;
YAC - acide malique ;
PVA - acide pyruvique ;
RBF - ribulose-1,5-bisphosphate ;
3-PGA - acide phosphoglycérique ;
PGA - phosphoglycéraldéhyde
Les réactions se déroulent comme suit. Dans les cellules mésophylles de la feuille, la carboxylation du PEP par les enzymes PEP-carboxylase se produit avec formation d'acide oxaloacétique PAA : PEP + CO 2 + H 2 O → PEP Cette substance instable est ici stabilisée par le NADPH avec formation d'acide malique (malate), un composé à 4 carbones : PIKE + NADPH → YAK. Dans un certain nombre de plantes, au lieu de l'AA, il se forme de l'acide aspartique (AA), qui nécessite de l'ammoniac dans les cellules : PIKE + NH 4 + NADPH → AA. Ces deux acides ne peuvent pas être formés dans la même plante, c'est pourquoi ces plantes sont appelées malate ou aspartate. La deuxième étape est que le malate passe dans les cellules tapissant le faisceau vasculaire, où, suite à sa décarboxylation oxydative, le CO 2 et le NADPH. sont formés pour le cycle Calvin. Le pyruvate (PVC) résultant retourne aux cellules du mésophylle et est phosphorylé en phosphoénolpyruvate, le principal accepteur de CO 2 , et la voie C4 est fermée. Le CO 2 libéré va à la deuxième carboxylation du RDP du cycle de Calvin avec formation de glucides dans les muqueuses cellulaires. La double carboxylation assure l'absorption du CO 2 de manière plus intensive que dans la voie C 3, c'est pourquoi elle est dite coopérative. La double carboxylation est beaucoup plus efficace et permet une photosynthèse intense avec une teneur réduite en CO 2. Ainsi, si chez les plantes C 3 la photosynthèse diminue en dessous de 0,03 %, alors chez les plantes C 4 elle se produit pleinement à 0,008 %. L'émergence d'une voie alternative pour le C 4 chez les plantes est considérée comme un processus d'évolution d'adaptation à une teneur réduite en CO 2 dans l'atmosphère et à une augmentation de l'O 2, due aux plantes elles-mêmes, que pendant la période de leur émergence. . Ces plantes présentent donc de grands avantages par rapport à la voie C 3 .
Le parcours C4 tire son nom du fait que dans la phase sombre, le principal produit de la fixation du CO2 est dans ce cas un composé organique comportant non pas trois, mais quatre atomes de carbone (acide oxaloacétique). Les plantes tropicales des pays chauds, par exemple les broméliacées, possèdent ce type de photosynthèse. On constate depuis longtemps que ces plantes absorbent bien mieux le CO2 que les plantes C3. Dans la structure anatomique des feuilles des plantes C4, à côté des chloroplastes ordinaires normaux, autour des faisceaux vasculaires se trouvent un type particulier de chloroplastes très denses, presque sans thylakoïdes, mais remplis d'amidon. Ces chloroplastes sont appelés chloroplastes pariétaux.
Dans les chloroplastes ordinaires des plantes C4, comme prévu, la phase légère de la photosynthèse se produit et la fixation du CO2 se produit également, mais en même temps de l'acide oxaloacétique se forme. Cet acide oxaloacétique est transformé en acide malique, qui pénètre dans les chloroplastes pariétaux, où il est immédiatement décomposé avec dégagement de CO2. Et puis tout se passe comme avec les installations C3 normales. Dans ce cas, la concentration de CO2 dans les chloroplastes pariétaux devient ainsi nettement plus élevée que dans les plantes C3, et la disposition très dense de ces chloroplastes garantit que presque aucun oxygène ne leur parvient et qu'il n'y a pas d'espaces intercellulaires ; Par conséquent, comme il n’y a pas d’oxygène et qu’il y a autant de dioxyde de carbone que l’on souhaite, la photorespiration ne se produit pas.
Ainsi, dans les plantes C4, la fixation du CO2 s'effectue plus efficacement sous forme d'autres composés, et la formation de sucres se produit dans des chloroplastes spéciaux, entraînant une réduction de l'intensité de la photorespiration et des pertes associées.
Les plantes C4 peuvent fermer leurs stomates sous l’effet de la chaleur et ne pas perdre cette précieuse humidité. Ils contiennent généralement suffisamment de CO2 accumulé sous forme d’acide malique.
27. Photorespiration : réactions biochimiques, leur localisation. Rôle physiologique de la photorespiration.
Photorespiration est un processus activé par la lumière de libération de CO2 et d’absorption d’O2. Le principal produit de la photorespiration est l’acide glycolique. La photorespiration augmente avec une faible teneur en CO2 et une concentration élevée en O2 dans l'air. Dans ces conditions, la chloroplaste ribulose disphate carboxylase catalyse non pas la carboxylation du ribulose-1,5-diphosphate, mais son clivage en acides 3-phosphoglycérique et 2-phosphoglycolique. Ce dernier est déphosphorylé pour former de l'acide glycolique.
L'acide glycolique passe du chloroplaste au peroxysome, où il est oxydé en acide glyoxylique. L'acide glyoxylique est aminé pour former la glycine. La glycine est transportée vers la mitochondrie, où la sérine est synthétisée à partir de deux molécules de glycine et où le CO2 est libéré.
La sérine peut pénétrer dans le peroxysome et transférer le groupe amino à l'acide pyruvique pour former l'alanine, et elle-même est convertie en acide hydroxypyruvique. Ce dernier, avec la participation du NADPH, est réduit en acide glycérique. Il passe dans les chloroplastes, où il entre dans le cycle de Calvin
Dans les usines C4 Le dioxyde de carbone de type 2 libéré lors de la photorespiration réagit dans les cellules du mésophylle avec l'acide phosphoénolpyruvique pour former des acides oxaloacétique et malique. L'acide malique passe dans les cellules de la gaine, où il sert de donneur de CO2. Plantes à voie C3 caractérisé par une haute intensité de photorespiration. L'acide phosphoglycolique se décompose par une série de transformations, libérant du CO2. Ainsi, lors de la photorespiration, une partie des produits intermédiaires de la photosynthèse est perdue du fait du dégagement de CO2. Les réactions d'oxydation et de carboxylation entrent en compétition, et la mise en œuvre de la fonction carboxylase ou oxygénase dépend de la teneur en O2 et CO2.
La photorespiration réduit l'efficacité de la photosynthèse et entraîne des pertes de carbone assimilé, mais a une certaine signification synthétique. Dans les premiers stades de la vie, lorsqu'il y avait peu d'oxygène dans l'atmosphère, le rubisco occupait une position clé dans la photosynthèse et sa fonction oxygénase ne posait pas de problèmes. À mesure que la teneur en oxygène augmentait, les pertes dues à la photorespiration augmentaient et un certain nombre de plantes développaient des mécanismes pour l'apport actif de dioxyde de carbone rubisco au lieu de travail (voir photosynthèse C4 et CAM), augmentant ainsi la part de son activité carboxylase à 100 %. .
La photosynthèse est le processus de transformation de l’énergie solaire absorbée par une plante en énergie chimique de composés organiques. Du 4 au voie de photosynthèse ou cycle Hatch-Slack.
Les scientifiques australiens Hatch et Slack ont décrit la voie C4 de la photosynthèse, caractéristique des plantes tropicales et subtropicales (canne à sucre, maïs…). Les feuilles de ces plantes contiennent deux types de chloroplastes : les chloroplastes ordinaires dans les cellules du mésophylle et les gros chloroplastes qui n'ont pas de grana ni de photosystème II dans les cellules de la gaine entourant les faisceaux vasculaires.
Dans le cytoplasme des cellules du mésophylle, il ajoute du CO 2 à l'acide pyruvique, formant de l'acide oxaloacétique. Il est transporté vers les chloroplastes, où il est réduit en acide malique avec la participation du NADPH. En présence d'ions ammonium, l'acide oxaloacétique est converti en acide aspartique. Les acides malique et (ou) aspartique passent dans les chloroplastes des cellules de la gaine et sont réduits en acide pyruvique et CO 2 . Le CO 2 est inclus dans le cycle de Calvin et l'acide pyruvique est transféré aux cellules du mésophylle, où il est converti en acide pyruvique. Ce mécanisme permet aux plantes de réaliser la photosynthèse lorsque les stomates sont fermés en raison de la température élevée. De plus, les produits du cycle de Calvin se forment dans les chloroplastes des cellules de la gaine entourant les faisceaux vasculaires. Cela favorise l'écoulement rapide des photoassimilats et augmente ainsi l'intensité de la photosynthèse.
Lipides : classification et leur rôle dans la plante.
Les lipides sont des substances dont la structure chimique est assez complexe. Ils comprennent également le carbone, l'oxygène et l'hydrogène, mais certains groupes de lipides peuvent inclure le phosphore, le soufre et l'azote (phosphatides, pigments). Tous les lipides sont hydrophobes, c'est-à-dire ne pas dissoudre dans l'eau. Les fonctions des lipides varient en fonction de leur structure chimique. Les lipides ne sont pas des biopolymères.
Les lipides sont classés en 5 grands groupes en fonction de leur fonction et de leur complexité structurelle : Les graisses - le groupe de lipides le plus facilement synthétisé. D'un point de vue chimique, il s'agit d'esters d'acides gras et de glycérol. Les principales fonctions des graisses sont l'énergie, la construction et le stockage. Les cires sont des substances grasses, solides à température ambiante. Selon leur structure chimique, ce sont des esters entre les acides gras et les alcools monohydriques de haut poids moléculaire de la série grasse. Les phosphatides - qui comprennent les glycérophosphatides, les lécithines et les céphalines - sont des molécules d'esters de glycérol, d'acides gras. et l'acide phosphorique. Ces substances font partie des graisses de stockage et les protègent du rancissement. La fonction principale des phosphatides est le stockage.
Les pigments (chlorophylles et caroténoïdes) constituent un groupe particulier de lipides à structure complexe, qui comprend également des radicaux azotés. Les pigments comprennent deux groupes de substances : les chlorophylles et les caroténoïdes.
La fonction principale des pigments est de participer à la phase énergétique (lumière) de la photosynthèse. - ce sont des dérivés d'un composé hétérocyclique complexe . Ce groupe de composés comprend les alcools de haut poids moléculaire (stérols) et leurs esters (stérides). Le stéroïde le plus connu est l'ergostérol, à partir duquel la vitamine D est obtenue industriellement.
Fonction principale des stéroïdes - construction (participer à la composition des membranes).
UDC 581.1:577.1
ASPECTS ÉVOLUTIONNAIRES DE LA PHOTOSYNTHÈSE C4
V.V. Ivanishchev
Les questions liées à la diversité des manifestations de l'assimilation photosynthétique du dioxyde de carbone chez les plantes et aux aspects évolutifs de la photosynthèse de type C4 sont prises en compte. Il est démontré qu'au cours des quinze dernières années, des données expérimentales ont été obtenues qui modifient considérablement nos idées sur la manière dont la photosynthèse C4 se produit et sur la mise en œuvre de ses propriétés particulières dans un certain nombre de plantes typiques C3. Les perspectives d'utilisation de ces connaissances pour développer des stratégies visant à améliorer l'efficacité de la photosynthèse des plantes C¡ afin d'augmenter leur productivité sont discutées.
Mots clés : plantes, assimilation du carbone inorganique, photosynthèse C3-C4, photosynthèse C4, aspects évolutifs, productivité végétale.
Introduction
La photosynthèse est la principale voie métabolique par laquelle le carbone inorganique est converti en composés organiques sur notre planète. L'étude des mécanismes du processus photosynthétique a conduit à la formation d'idées sur les voies biochimiques d'assimilation du carbone inorganique de types C3 et C4. Après la découverte de la réaction oxygénase Rubisco f-ribulose-1,5-bisphosphate carboxylase oxygénase, EC 4.1.1.39), il était d'usage de parler de photorespiration comme d'un processus parallèle. Spécialistes individuels, incl. L'auteur, qui a apporté une contribution significative à la compréhension de la biochimie de la photorespiration, a commencé à identifier ce qu'on appelle la photosynthèse C2. Découverte des caractéristiques du transport (pas dans les mitochondries et les peroxysomes de la même cellule) et du métabolisme du phosphoglycolate (formé lors de l'oxydation du D-ribulose-1,5-bisphosphate avec l'oxygène), mais de son transfert et de son métabolisme dans les cellules de la gaine avec suivi « capter » le CO2 libéré par les chloroplastes. Ces cellules ont permis de formuler des idées sur la photosynthèse du C2. Dans ce cas, même la réaction oxygénase de Rubisco est considérée comme un fournisseur de CO2, de sorte que l'efficacité globale de la photosynthèse n'est au moins pas fortement réduite.
Des études actives sur les plantes dont la vie est confinée à des conditions environnementales spécifiques au cours des trois dernières décennies ont révélé certaines variations dans la manifestation des différentes voies du métabolisme du carbone au cours de la photosynthèse. Dans le même temps, une plasticité morphologique et biochimique élevée des plantes C3 et C4 a été révélée. De plus, divers mécanismes ont été découverts grâce auxquels les plantes de type C3 présentaient un certain nombre de propriétés caractéristiques des plantes de type C4.
Dans les conditions écologiques spécifiques de salinité et de sécheresse de l'Asie centrale, des plantes ont été découvertes possédant une voie photosynthétique de type C4 sans l'anatomie classique des feuilles de Kranz. De plus, grâce à une autre découverte, la frontière nette entre les deux principaux types de plantes en termes de mécanisme de fixation du CO2 est effacée. Il a été démontré que dans les cellules entourant la nervure centrale du système conducteur des feuilles de riz (une plante typique C3), des enzymes clés caractéristiques de la photosynthèse de type C4 sont présentes.
Tout cela rend nécessaire de généraliser les résultats obtenus afin de déterminer d'autres orientations de recherche dans ce domaine scientifique et les moyens possibles d'augmenter l'efficacité de la photosynthèse (et la productivité) des plantes classiques C3, qui constituent la majorité des plus importantes. les cultures agricoles.
Caractéristiques anatomiques, morphologiques et plasticité
photosynthèse C4
Les plantes dotées de photosynthèse de type C4 sont connues pour avoir tout un complexe de caractéristiques. Ils confèrent aux plantes un certain nombre de caractéristiques pratiques importantes - une plus grande résistance à certains facteurs environnementaux (températures élevées, sécheresse), une consommation d'eau plus économique, une productivité plus élevée avec une consommation moindre d'azote, ce qui explique l'intérêt économique des producteurs à cultiver de telles cultures. Les propriétés caractéristiques de la structure des feuilles fournissent tout d'abord aux plantes C4 un mécanisme de concentration de CO2 qui leur permet d'éviter la perte de dioxyde de carbone et ainsi de réduire la photorespiration, grâce à quoi l'efficacité de la photosynthèse chez les plantes C3 peut diminuer jusqu'à à 40%, surtout dans des conditions de températures élevées et de sécheresse.
Chez les plantes C3, on peut observer deux types de cellules physiologiquement et spatialement séparés, à savoir : un groupe est plus ou moins uniformément réparti et forme le mésophylle, l'autre partie des cellules est concentrée autour des faisceaux vasculaires. Cette photo les fait ressembler à des plantes C4. Cependant, biochimiquement, ces cellules sont identiques.
D'autre part, certaines plantes C3 présentent les caractéristiques des plantes C4 en l'absence de deux types de cellules anatomiquement sensiblement différents, ce qui est cohérent avec les idées classiques de la voie de fixation du dioxyde de carbone C4. Malgré cela, la photosynthèse C4 se caractérise par un certain nombre de caractéristiques fonctionnelles strictement spécifiques. Comprendre les transformations évolutives dans cet aspect
est basé sur des idées sur les changements dans les caractéristiques cellulaires et leur prédétermination génétique.
Dans la plupart des plantes C4, le fonctionnement de la PEP carboxylase et le cycle de Calvin sont spatialement séparés entre ces deux types de cellules. La densité élevée des veines des faisceaux vasculaires conduit au fait que le rapport entre les volumes de tissus formés par les cellules du mésophylle et la muqueuse des faisceaux vasculaires est d'environ 1:1.
L'étude de la formation de ces types de cellules montre qu'au cours de l'ontogenèse, les couches internes des cellules peuvent se différencier des cellules du procambium, et les couches externes des cellules du méristème. L'analyse phylogénétique montre que les plantes C4 ont émergé via plusieurs voies évolutives différentes. Dans ce cas, les parois des cellules tapissant les faisceaux vasculaires peuvent contenir de la subérine, ce qui limite la diffusion du CO2 au-delà des limites de ces cellules. Dans le même temps, une couche similaire de subérine se trouve également dans certaines plantes C3 sur les parois externes des couches internes de la muqueuse des faisceaux vasculaires. Ainsi, aucune caractéristique spécifique aux cellules foliaires des plantes C4 n’a encore été découverte.
Un autre point important est la distance entre les deux types de cellules afin d'assurer le taux d'échange maximal de métabolites entre elles. Ceci explique que le nombre de couches de la cage de revêtement soit limité à une ou deux. Dans le même temps, une augmentation de l'efficacité du processus de photosynthèse est obtenue, d'autre part, par un petit nombre de cellules mésophylles entre les cellules de la gaine, dont le nombre peut aller jusqu'à quatre chez certaines espèces. Dans le même temps, le nombre de cellules entre les faisceaux vasculaires est généralement faible. Pour réduire la distance entre les deux types de cellules, on retrouve parfois des cellules achlorophylles dans les feuilles.
Dans le même temps, l'étude de diverses plantes dicotylédones C4 a montré que, par rapport aux plantes typiques C3, bon nombre de leurs caractéristiques anatomiques et morphologiques se sont révélées similaires. Cela s'applique à des indicateurs tels que la densité des nervures pour 1 mm2 de surface des feuilles, l'épaisseur des feuilles et le pourcentage d'espace intercellulaire. D’autres indicateurs avaient des limites de variabilité plus ou moins larges, ou étaient décalés vers des valeurs plus élevées ou plus basses.
La troisième circonstance importante caractéristique des plantes C4 est un grand compartiment qui assure la liaison maximale possible (sans perte) du CO2 libéré lors de la décarboxylation du composé à quatre carbones. Ceci explique la présence d'un nombre accru de chloroplastes présents dans les cellules de la gaine. L'excès du volume des cellules de la gaine sur le volume des cellules du mésophylle est obtenu de plusieurs manières, à savoir : en augmentant le nombre de chloroplastes dans les cellules de la gaine, en augmentant le nombre de cellules de la gaine, ou
Le nombre de faisceaux conducteurs ou les changements dans le diamètre des veines. Les indicateurs décrits sont interdépendants et, en partie, déterminés par les conditions environnementales, qui fournissent avant tout les paramètres nécessaires à l'échange d'eau et les indicateurs associés, tels que la pression hydraulique, le débit et le mouvement de l'eau dans l'usine, etc. . Tout cela détermine la diversité des manifestations anatomiques du syndrome C4.
La dernière caractéristique des plantes C4 est que les chloroplastes des cellules de la gaine du faisceau vasculaire et les cellules du mésophylle diffèrent non seulement en nombre, mais aussi morphologiquement et fonctionnellement. Dans le même temps, un grand nombre de chloroplastes dans les cellules de la gaine est caractéristique non seulement des plantes C4, mais également des plantes à photosynthèse dite C2, dans lesquelles la glycine formée lors de la photorespiration est transférée des cellules du mésophylle aux cellules de la gaine, où le CO2 libéré au cours de son métabolisme est ensuite capturé par les chloroplastes et utilisé dans le cycle de Calvin. Ces particularités de la mise en œuvre de ce type de photosynthèse sont considérées par les chercheurs comme l'une des manières par lesquelles l'évolution des mécanismes d'assimilation photosynthétique du carbone inorganique peut être représentée sous la forme de la séquence : C3^C2^C4. D'autres caractéristiques du syndrome C4 incluent des caractéristiques de la répartition des organelles telles que les mitochondries et les peroxysomes entre différentes cellules des feuilles des plantes, ainsi que des caractéristiques de l'ultrastructure et des propriétés photochimiques des chloroplastes.
Les indicateurs décrits ci-dessus sont très flexibles. En fonction des conditions environnementales, ils peuvent changer de manière significative, grâce à quoi les plantes dotées de la voie de photosynthèse C3 peuvent acquérir des traits caractéristiques de la voie C4. Dans le même temps, la variété des changements anatomiques et morphologiques possibles dans divers traits des plantes C4 leur permet de maintenir leurs caractéristiques physiologiques et biochimiques au niveau requis. Cependant, en général, la plasticité des plantes C3 dans cet aspect est mieux évaluée, malgré le fait que les données expérimentales dans ce domaine font clairement défaut.
Caractéristiques biochimiques des manifestations de la photosynthèse C4
dans les plantes supérieures
Les idées générales classiques sur la voie C4 pour la fixation du carbone inorganique incluent trois variantes principales dans l'utilisation du premier produit stable - un composé à quatre carbones (malate et/ou aspartate) comme source de CO2 pour le D-ribulose-1,5- bisphosphate carboxylase oxygénase (Rubisco) - enzyme clé du cycle de Calvin. Ces formes de plantes sont désignées comme NADP-
malik-enzyme, NAD-malik-enzyme et PEP-carboxykinase - du nom de l'enzyme clé qui assure la décarboxylation d'un composé à quatre carbones avec libération de CO2. Plus tard, 14 variétés de combinaisons anatomiques et biochimiques ont été découvertes dans les graminées de type C4. Ils différaient par des caractéristiques telles que la taille des cellules du mésophylle et des cellules tapissant les faisceaux vasculaires, le volume total de cellules de différents types, l'emplacement des chloroplastes dans les cellules, la présence du nombre de grana dans les chloroplastes, etc. avec des variations correspondantes dans la distribution cellulaire et l'ampleur de l'activité enzymatique.
Contrairement à ces herbes, seuls deux types de plantes sont connus chez les dicotylédones : le type NADP-malik-enzyme et le type NAD-malik-enzyme, tandis que la PEP carboxykinase est impliquée dans les voies secondaires du métabolisme des glucides chez un certain nombre d'espèces. De plus, les caractéristiques anatomiques et biochimiques de ces plantes sont également associées aux caractéristiques environnementales de leur croissance et de leur développement. Par exemple, l’étude a permis d’identifier cinq variétés d’anatomie des feuilles de Kranz chez les représentants C4 de la famille des Chénopodiacées.
Une étude des caractéristiques biochimiques des plantes dicotylédones C4 a montré qu'en général, les indicateurs décrits étaient similaires à ceux caractéristiques d'autres espèces C4. A titre caractéristique, il convient de noter encore une fois que la PEP carboxykinase n'apporte pas de contribution significative au métabolisme des acides dicarboxyliques, contrairement aux graminées monocotylédones. En revanche, la présence simultanée de deux enzymes décarboxylantes dans une espèce (enzyme NAD-malik + PEP carboxykinase ou enzyme NADP-malik + PEP carboxykinase) reste largement floue. Cela peut s'expliquer en partie, par exemple, par la nécessité de synthétiser du PEP (pour la voie du shikimate), d'économiser de l'ATP pour soutenir d'autres processus, etc. .
En conséquence, il a été constaté que parmi les plantes de type C4, les espèces avec une anatomie de feuille atriplicoïde (caractéristique des représentants du genre Atriplex) et une biochimie enzymatique NADP-malik prédominent.
Caractéristiques des centrales « intermédiaires » C3-C4
L'étude de divers représentants du règne végétal a permis de parler d'un groupe photosynthétique spécifique, qui a commencé à être désigné comme plantes C3-C4. Les traits caractéristiques de ce groupe (intermédiaires C3-C4) sont les caractéristiques uniques de l'échange gazeux dans les feuilles, qui montrent un faible point de compensation du CO2 et sa dépendance curviligne à la concentration en oxygène dans l'environnement. Cela est dû à la compartimentation particulière (lieu d'apparition) de la photorespiration dans les cellules sans modification du métabolisme photosynthétique du carbone. Majorité
les représentants des intermédiaires ont une photosynthèse de type C3 et leur principale caractéristique est la présence d'une photorespiration réduite
La structure anatomique des feuilles des intermédiaires C3-C4 les distingue des plantes typiques C3 et C4. Contrairement aux plantes C4, chez les plantes intermédiaires, les cellules du mésophylle ne sont pas concentrées autour des cellules qui tapissent les faisceaux vasculaires et sont plus lâchement situées dans la structure des feuilles. Dans le même temps, les cellules intermédiaires contiennent un grand nombre d'organites, dont beaucoup sont situés de manière centripète (vers le centre) dans les cellules de la gaine. Le nombre de mitochondries et de peroxysomes est ici quatre fois plus élevé que dans les cellules mésophylles, et les mitochondries sont deux fois plus grandes. Dans le même temps, le nombre de plasmodesmes entre les cellules de la gaine et du mésophylle est beaucoup plus important que chez les plantes C3 et se rapproche du chiffre caractéristique du type C4. Cela garantit un échange rapide de métabolites entre les différentes cellules des feuilles des plantes.
L'enzyme clé responsable de la libération de CO2, la glycine décarboxylase, est principalement localisée dans les cellules de la gaine. Les particularités de la disposition relative des différents organites dans les cellules permettent aux plantes C3 de capter jusqu'à 50 % du CO2 formé (dans les réactions de conversion de la glycine), tandis que pour les intermédiaires C3-C4 cette valeur peut atteindre 75 %.
En conséquence, ces caractéristiques permettent aux intermédiaires de présenter de meilleures caractéristiques d’efficacité d’assimilation photosynthétique du CO2, en particulier à des températures élevées, dans l’utilisation de l’eau, etc., qui sont décrites avec succès à l’aide d’un modèle mathématique.
L'étude des caractéristiques de la biochimie a montré que les intermédiaires n'ont pas la distribution spécifique typique des enzymes clés (PEP carboxylase et Rubisco) dans différentes cellules, comme c'est le cas dans les plantes avec photosynthèse de type C4, ils montrent l'inefficacité du composants existants des systèmes enzymatiques de type C4, et se caractérisent par une faible activité des enzymes associées à l'échange d'acides C4-dicarboxyliques (PEP carboxylase, NADP malate déshydrogénase, enzyme malique NADP, pyruvate orthophosphate dikinase). Dans ce cas, des quantités importantes de carbone marqué provenant du 14CO2 assimilé entrent dans la composition de la glycine, de la sérine et surtout du fumarate. Dans le même temps, les avantages de certaines plantes C3-C4 sont dus uniquement à la présence d'un échange actif de glycine entre les cellules des feuilles, comme indiqué ci-dessus. Cependant, le métabolisme du carbone actif doit s’accompagner d’un bilan d’azote actif, mais le tableau de ce processus reste inconnu.
Photosynthèse C4 sans anatomie des feuilles de Kranz
La photosynthèse C4, due à la présence de l'anatomie de Kranz de la feuille, suggère que la paroi cellulaire entre la gaine et les cellules du mésophylle empêche la perte de CO2 libéré lors de la décarboxylation du métabolite C4. Cependant, l'étude de la diversité végétale associée à l'existence dans diverses niches écologiques a permis de découvrir des plantes d'Asie centrale dans les feuilles desquelles de nombreuses caractéristiques de la photosynthèse C4 sont réalisées au sein d'une seule cellule sans changement fondamental dans l'anatomie des feuilles. Parallèlement, les données connues sur les mécanismes de concentration du CO2 dans les plantes aquatiques, ainsi que la présence d'un cycle photosynthétique C4 dans certaines d'entre elles au sein d'une cellule individuelle, sont largement déterminées par les conditions d'existence en milieu aquatique.
En revanche, chez les plantes terrestres, il doit exister une frontière physique (paroi cellulaire épaisse, membrane, enveloppe) séparant les cellules où se produit l'assimilation photosynthétique du dioxyde de carbone et les cellules où fonctionne le cycle de Calvin (Fig.).
Schéma simplifié de compartimentation de la C^-photosynthèse (deux types de cellules sont représentés, entre lesquelles l'assimilation du carbone inorganique et son incorporation sont spatialement séparées
en produits du cycle de Calvin)
Les plantes terrestres de la famille CHnporoShasvav, représentées par plus de 1 300 espèces, sont souvent résistantes à la sécheresse et à la salinité, ce qui est largement assuré par la présence de photosynthèse de type C4. Parmi les représentants de cette famille, des espèces ont été découvertes dans lesquelles la photosynthèse C4 est réalisée au sein d'une cellule sans
présence de l'anatomie classique des feuilles de Kranz. Il s'agit de Bienertia cycloptera, B. sinuspersici, Suaeda aralocaspica. Ils poussent dans des conditions de salinité et/ou de manque d’eau. Dans le même temps, les tailles des plantes diffèrent considérablement, à savoir : si la hauteur de B. cycloptera et Suaeda aralocaspica est de 20 ... 50 cm, alors les plantes de B. sinuspersici peuvent atteindre 130 ... 160 cm.
Chez ces espèces, deux types de répartition topographique des organites au sein des cellules ont été trouvés. Ainsi, dans les cellules de X aralocaspica, on distingue les parties distale et proximale. Dans la partie distale, sont localisés les chloroplastes agranaux, dans lesquels l'amidon n'est pas synthétisé, mais il existe une enzyme - pyruvate orthophosphate dikinase pour la synthèse du PEP, et ici (dans cette partie de la cellule) le CO2 est fixé avec la formation d'un Produit C4. Dans la partie proximale de la cellule, les chloroplastes avec les enzymes du cycle de Calvin et l'enzyme malik sont concentrés, grâce à quoi le malate est converti en pyruvate avec libération de CO2 pour son inclusion dans les produits de réaction du cycle de Calvin avec la participation de Rubisco et d'autres enzymes.
Dans les cellules des espèces du genre Bienertia, on distingue un compartiment central et périphérique. Dans la partie périphérique, il y a des chloroplastes avec un petit nombre de grains, presque pas de mitochondries, mais les enzymes clés de la voie C4 sont présentes - la pyruvate orthophosphate dikinase et la PEP carboxylase, qui assurent respectivement la synthèse du PEP et la fixation du dioxyde de carbone. Les mitochondries et les chloroplastes avec une structure granulaire développée, une grande quantité de Rubisco et d'autres enzymes du cycle de Calvin sont concentrés dans la partie centrale de la cellule. Dans ce cas, le compartiment central est entouré d'une vacuole (vacuoles) pour éviter la perte de CO2.
Dans le même temps, les espèces décrites ont différents ratios d'activités d'enzymes clés importantes pour la mise en œuvre des voies C3 et C4, ce qui les distingue des véritables espèces C4.
Une autre découverte intéressante est la découverte d'un type de photosynthèse de type C4 dans les tiges et les pétioles du tabac, ainsi que dans les nervures médianes des feuilles d'Arabidopsis (Arabidopsis LaNapa L.) - représentants des dicotylédones. Ces particularités sont liées à la présence d'une activité élevée de toutes les enzymes clés du syndrome C4, incl. la présence simultanée d'une activité élevée de toutes les enzymes décarboxylantes (enzymes maliques NAD et NADP, ainsi que PEP carboxykinase).
Plus tard, une découverte similaire a été faite pour les représentants des monocotylédones. Ainsi, dans les cellules situées autour de la veine centrale du faisceau vasculaire de la feuille d'une plante C3 typique - le riz, une activité accrue similaire des enzymes ci-dessus a été trouvée. L'utilisation de l'analyse par immunoblot par les auteurs donne le résultat obtenu
les résultats ont une importance significative en tant que base de données probantes. Il a été démontré que l'activité de Rubisco dans les cellules de cette partie est deux fois inférieure à celle des autres parties du limbe. L'activité de la PEP carboxylase est augmentée d'environ 30 %. Dans le même temps, l’activité de chacune des enzymes décarboxylantes (enzymes maliques NAD et NADP, ainsi que PEP carboxykinase) dans la région de la nervure médiane de la feuille était six fois ou plus supérieure. L’activité d’une autre enzyme importante, la pyruvate orthophosphate dikinase, était 7,5 fois supérieure à celle des cellules du limbe des feuilles.
D'une part, au cours de la discussion, les chercheurs interprètent ces données comme l'une des voies naturelles (naturelles) de l'évolution et de l'émergence possibles de la photosynthèse C4, d'autre part, les auteurs considèrent une telle image comme une opportunité de manipulation humaine pour "améliorer" la photosynthèse des plantes C3 (sur les discussions en cours depuis le milieu des années 70 du siècle dernier). Cela s'applique tout d'abord à une culture agricole d'importance mondiale comme le riz.
Dans le même temps, les auteurs de l'étude ont consacré une partie importante de leurs travaux à l'étude des particularités du système pigmentaire chloroplastique, qui fournit de l'énergie pour la synthèse de composés organiques lors de la photosynthèse.
La photosynthèse C4 à la lumière de l'évolution
L'apparition de plantes avec une photosynthèse de type C4 au cours de l'évolution, comme le pensent de nombreux auteurs, s'est produite plusieurs dizaines de fois de manière indépendante. La majeure partie de ces plantes est aujourd'hui représentée par des graminées (environ 4 600 espèces) et des carex (1 600 espèces), tandis que chez les dicotylédones, les plantes de type C4 sont représentées par environ 1 600 espèces, ce qui suggère une origine évolutive polyphylétique du syndrome C4. Dans le même temps, l'émergence évolutive d'une nouvelle voie d'assimilation photosynthétique du CO2 aurait dû inclure un certain nombre d'étapes affectant le développement de l'anatomie des feuilles de Kranz, la création d'une pompe à CO2 pour concentrer le dioxyde de carbone, la localisation et la formation du cycle C4. , des changements dans l'expression des gènes avec une optimisation supplémentaire des propriétés enzymatiques.
Une image théorique assez claire et simple de l'émergence de la photosynthèse C4 soulève bien entendu également des questions concernant les modifications des voies métaboliques d'autres éléments chimiques et, en premier lieu, de l'azote et du soufre. Par conséquent, l'apparition des plantes céréalières atteintes du syndrome C4 pourrait avoir eu lieu il y a assez longtemps - il y a environ 30 millions d'années, tandis que l'apparition des dicotylédones présentant ce type de photosynthèse est estimée à 20 millions d'années.
L’émergence de la voie photosynthétique C4 était auparavant envisagée à travers l’apparition d’intermédiaires C3-C4 comme formes de transition. L'étude de la variété des caractéristiques de la disposition des organites dans les cellules du mésophylle et de la muqueuse des faisceaux vasculaires, ainsi que d'un certain nombre d'enzymes, a montré que le mécanisme clé de l'apparition du syndrome C4 semble être la redistribution entre différentes cellules de glycine décarboxylase, dont l'activité était fortement réduite ou perdue dans les cellules du mésophylle. Une preuve indirecte de cette voie est que l’activité de cette enzyme dans les plantes C4 est présente uniquement dans les cellules de la gaine. Dans le même temps, on ne sait pas pourquoi, chez de telles espèces C3-C4, en présence d'une « navette » (transfert) de glycine d'un type de cellule à un autre suivie d'une décarboxylation, les avantages théoriquement prédits en termes de taux d'assimilation du CO2 et l'efficacité de l'utilisation de l'eau et de l'azote ne se manifeste pas dans des conditions de concentrations normales de dioxyde de carbone.
L'examen des possibilités de modifications anatomiques de la structure des feuilles montre que chaque caractéristique caractéristique du syndrome C4 peut changer indépendamment chez les plantes C3 (taille des cellules de la mésophylle et de la gaine, épaisseur des feuilles, distance entre les nervures des faisceaux vasculaires), tandis que pour la photosynthèse C4. de tels changements est important. De ce point de vue, de nombreux chercheurs envisagent la transition vers les plantes C4 par l’émergence de la photosynthèse dite C2, en se concentrant sur le transport de la glycine et sa décarboxylation dans les cellules de la gaine.
Ainsi, l’origine polyphylétique du syndrome C4 n’est devenue possible que grâce à l’émergence de nombreux traits caractéristiques, coordonnés lors de la sélection naturelle dans des conditions environnementales changeantes.
Perspectives de création de mécanismes de concentration
CO2 dans les usines C3
Des recherches théoriques et un certain nombre de données expérimentales ont permis d'évoquer la potentielle transformation génétique des plantes C3, grâce à laquelle il serait possible de les améliorer (les rapprocher des plantes C4 en termes d'indicateurs photosynthétiques, de résistance aux facteurs environnementaux et productivité). Plus tard, la possibilité d'un mécanisme similaire fonctionnant uniquement avec la participation de l'oxaloacétate décarboxylase a été prouvée expérimentalement.
Actuellement, la solution au problème de l'augmentation de l'efficacité de la photosynthèse (et, par conséquent, de l'augmentation de la productivité) des plantes C3, qui représentent l'essentiel des cultures agricoles cultivées, est envisagée de plusieurs manières.
Dans le même temps, bon nombre des solutions proposées soulèvent des objections raisonnables.
Les travaux récents sont particulièrement intéressants à cet égard, qui sont consacrés non seulement à une analyse détaillée des caractéristiques de la manifestation du syndrome C4 pour comprendre l'évolution des voies photosynthétiques, mais également à la justification du déterminisme génétique de l'évolution du syndrome C3. plantes dans le type C4. Il est à noter que jusqu'à présent, les mécanismes génétiques qui ont assuré l'apparition chez les plantes d'une anatomie foliaire spécifique caractéristique de la photosynthèse C4 restent inconnus, et c'est le principal problème des tentatives de bio-ingénierie visant à conférer de nouvelles caractéristiques ou à transformer la plupart des plantes agricoles C3 les plus importantes. en type C4.
Les caractéristiques des mécanismes génétiques peuvent consister dans le fait que de nombreuses caractéristiques de la photosynthèse C4 sont apparemment réalisées à l'aide de nombreux gènes, dont la manifestation de l'activité a de petits effets, qui ne donnent que collectivement l'effet nécessaire. Dans le même temps, les voies du déterminisme génétique, comme le montrent les données sur diverses plantes C3-C4 et C4, peuvent être diverses, puisque chez les représentants de familles et de genres individuels, les formes transitionnelles de C3^C3-C4^C4 ont caractéristiques et propriétés particulières. À cet égard, il est d’abord nécessaire de suivre ces caractéristiques de transition dans les familles et genres de plantes qui sont taxonomiquement proches des représentants correspondants de cultures agricoles importantes.
Les problèmes rencontrés lors de la transformation d'usines C3 en plantes de type C4 sont décrits dans les travaux de Gowik et Westhoff. Les auteurs notent non seulement la nécessité de séparer les réactions impliquant la PEP carboxylase et la Rubisco dans différentes cellules, mais soulèvent également des questions sur la nécessité de modifier l'expression de différents gènes. Cela est dû, par exemple, au fait que la quantité de Rubisco dans les plantes C4 est plusieurs fois inférieure à celle des plantes C3. De plus, les mécanismes de régulation dans chaque type de cellule (mésophylle et gaine) diffèrent significativement. Ainsi, dans la plante C4 Flaveria trinervia, un élément cis-régulateur est présent dans les cellules du mésophylle, désigné sous le nom de MODULE D'EXPRESSION DE LA MÉSOPHYLLE1. Le même élément génétique est également présent dans les gènes des plantes de type C3 de ce genre Flaveria, mais il ne possède pas la spécificité de manifestation nécessaire. Une légère modification d'un tel élément suffit à l'expression spécifique du gène spécifié.
Contrairement à ce qui précède, l'expression spécifique (pour les cellules de la gaine du faisceau vasculaire) de l'un des gènes codant pour la petite sous-unité de Rubisco, FbRbcSl, est régulée principalement au niveau post-transcriptionnel. Il semble que la stabilité des transcrits de ce gène dans les cellules du mésophylle et de la gaine soit différente.
Ainsi, la transformation des plantes C3 en type C4 est largement associée à des changements dans la régulation de l'activité de divers gènes. Dans ce cas, le rôle clé semble être joué par les facteurs de transcription GOLDEN2-LIKE GLK1 et GLK2, caractéristiques de toutes les plantes terrestres. Chez la plante Arabidopsis, ils contrôlent l’expression de plus de 100 gènes, dont beaucoup sont associés au processus photosynthétique, tandis que chez le maïs (plantes C4), une expression spécifique des cellules de la gaine de ce groupe a été observée. Ces résultats ont amené les auteurs à conclure que ces protéines sont importantes pour la régulation des gènes dans la formation des cellules mésophylles et la muqueuse des faisceaux vasculaires. Cependant, d’autres étapes pourraient impliquer l’optimisation des quantités et des propriétés de nombreuses enzymes.
Ainsi, l'évolution de la photosynthèse apparaît très complexe et diversifiée, et les problèmes de bio-ingénierie et de contrôle de la productivité des plantes à l'aide de manipulations génétiques semblent avoir des perspectives de solution, compte tenu des connaissances accumulées sur les voies de régulation des caractéristiques morphologiques et biochimiques. important pour la manifestation d’une photosynthèse plus efficace de la voie C4.
Bibliographie
1. Heldt G.-W. Biochimie des plantes. M. : BINOM. Laboratoire des Connaissances, 2011. 471 p.
2. Tolbert N. E. Le cycle du carbone photosynthétique oxydatif C2 // Annu. Tour. Physiol végétal. Plante Mol. Biol. 1997. V. 48. P. 1-25.
3. Lundgren M.R., Osborne C.P., Christin P.-A. Déconstruire l'anatomie de Kranz pour comprendre l'évolution du C4 // J. Exp. Bot. 2014. V. 65 (13). P. 33573369.
4. Preuve de photosynthèse C4 sans anatomie de Kranz chez Bienertia cycloptera (Chenopodiaceae) / E.V. Voznesenskaya // Le Journal des Plantes. 2002. V. 31 (5). P. 649-662.
5. Sage R.F. La photosynthèse C4 dans les plantes terrestres ne nécessite pas l'anatomie de Kranz // TENDANCES en science végétale. 2002. V. 7 (7). P. 283-285.
6. Wickramage A.T. Photosynthèse unicellulaire C4. 2008. https://www.slideshare.net/anupathisalwickramage/single-cell-c4photosynthesis.
7. L'existence d'une photosynthèse de type gaine C-4 dans la nervure médiane du riz C-3 / W.J. Shen // Riz. 2016. V. 9. N° 20. DOI : 10.1186/s12284-016-0094-5.
8. Gowik U., Westholl P. Le chemin de la photosynthèse C3 à C4 // Plant Physiol. 2011. V. 155. P. 56-63.
9. Muhaidat R., Sage R.F., Dengler N.G. Diversité de l'anatomie et de la biochimie de Kranz chez les eudicots C4 // Amer. J. Bot. 2007. V.94 (3). P. 362-381.
10. La photosynthèse C2 génère des niveaux de CO2 foliaires environ 3 fois plus élevés chez l'espèce intermédiaire C3-C4 Flaveria pubescens / O. Keerberg //
Journal de botanique expérimentale. 2014. V. 65. Non. 13. P. 3649-3656. est ce que je:10.1093/jxb/eru239.
11. La glycine décarboxylase est confinée aux cellules de la gaine des feuilles des espèces intermédiaires C3-C4 / C.M. Hylton // Plante. 1988. V. 175. P. 452-459.
12. Sage R.F., Sage T.L., Kocacinar F. Photorespiration et évolution de la photosynthèse C4 // Revue annuelle de biologie végétale. 2012. V. 63. P. 19-47.
13. L'inférence phénotypique du paysage révèle de multiples voies évolutives vers la photosynthèse C4 / B.P. Williams // eLife 2. 2013. e00961. est ce que je.org/10.7554/eLife.00961.
14. Edwards J., Walker D. Photosynthèse des plantes C3 et C4 : mécanismes et régulation. M. : Mir, 1986. 590 p.
15. Monson R.K. Assimilation du CO2 dans les plantes intermédiaires C3-C4 // Photosynthèse : Physiologie et Métabolisme / R.C. Leegood, T.D. Sharkey, S. von Caemmerer (éd.). Éditeurs académiques Kluwer. 2000. P. 533-550.
16. Sage R.F. L'évolution de la photosynthèse C4 // Nouveau phytologue. 2004. V. 161. P. 341-370.
17. Hunt S., Smith A.M., Woolhouse H.W. Preuve d'un système dépendant de la lumière pour la réassimilation de la photorespiratoire qui n'inclut pas de cycle, chez l'espèce intermédiaire Moricandia arvensis // Planta. 1987. V. 171. P. 227-234.
18. von Caemmerer S. Un modèle d'assimilation photosynthétique et de discrimination des isotopes du carbone dans les feuilles de certains intermédiaires // Planta. 1989. V. 178. P. 463-474.
19. Ivanishchev V.V. Sur la détermination de l'efficacité de la carboxylation chez les plantes dotées d'un mécanisme de concentration du CO2 // Physiologie végétale. 1992. T. 39. N° 3. P. 437444.
20. Hibberd J.M., Quick W.P. Caractéristiques de la photosynthèse C4 dans les tiges et pétioles des plantes à fleurs C3 // Nature. 2002. V. 415 (6870). P. 451-454. est ce que je:10.1038/415451a.
21. Les décarboxylases acides en C4 nécessaires à la photosynthèse en C4 sont actives dans la veine médiane de l'espèce C4 Arabidopsis thaliana et sont importantes dans le métabolisme des sucres et des acides aminés / N.J. Brown // Plante J. 2010. V. 61(1). P. 122-133. est ce que je:10.1111/j.1365-313X.2009.04040.x.
22. Magomedov I.M. Sur l'histoire de la découverte de la photosynthèse C4. État actuel du problème // Progrès des sciences naturelles modernes. 2015. N° 1 (partie 6). pages 962 à 965.
23. Nasyrov Yu.S. Génétique de la photosynthèse et de la sélection. M. : Connaissance, 1982. 64 p.
24. Abdullaev A., Gorenkova L.G., Ivanishchev V.V. Sur la distribution de certaines enzymes du métabolisme de l'acide C4 dans les chloroplastes de seigle // Physiologie végétale. 1989. T. 36. N° 4. P. 665-668.
25. Ivanishchev V.V. Importance biologique du métabolisme de l'oxaloacétate dans les chloroplastes des plantes C3 // Physiologie végétale. 1997. T. 44. N° 3. P. 462-470.
26. Meyer V., Griffiths H. Origines et diversité des mécanismes eucariotiques de concentration de CO2 : leçons pour l'avenir // J. Exp. Bot. 2013. V. 64. N° 3. P. 769-786.
27. Ivanishchev V.V. Problèmes d'assimilation photosynthétique du carbone inorganique par les plantes supérieures // Bulletin du GOU DPO TO "IPK ET PPRO TO". Espace éducatif de Toula. 2017. N° 3 (sous presse).
28. Evolution et fonction d'un module cis-régulateur pour l'expression génique spécifique du mésophylle dans le dicotylédone C4 Flaveria trinervia / M. Akyildiz // Cellule végétale. 2007. V. 19. P. 3391-3402.
29. Patel M., Siegel A.J., Berry JO. Les régions non traduites de l'ARNm FbRbcS1 médient l'expression génique spécifique des cellules de la gaine du faisceau dans les feuilles d'une plante C4 // J. Biol Chem. 2006. V. 281. P. 25485-25491.
30. Waters M.T., Langdale J.A. La fabrication d'un chloroplaste // EMBO J. 2009. V. 28. P. 2861-2873.
Ivanishchev Viktor Vasilievich, docteur en biologie. Sciences, Arts. scientifique employé, gérant département, [email protégé], Russie, Toula, Université pédagogique d'État de Toula. L.N. Tolstoï
ASPECTS ÉVOLUTIONNAIRES DE C4-PHOTOSYNTHÈSE V.V. Ivanishchev
La revue traite de la variété des manifestations de l'assimilation photosynthétique du dioxyde de carbone chez les plantes et des aspects évolutifs de la photosynthèse de type C4. Il est démontré qu'au cours de la dernière décennie et demie, des données expérimentales ont été obtenues qui modifient considérablement notre compréhension des modes d'apparition de la photosynthèse C4 et la réalisation de ses propriétés spécifiques dans un certain nombre de plantes typiques C3. Les perspectives d'application de ces connaissances pour développer une stratégie d'amélioration de l'efficacité de la photosynthèse des plantes C3 sont discutées afin d'augmenter leur productivité.
Mots clés : plantes, assimilation du carbone inorganique, photosynthèse C3-C4, photosynthèse C4, aspects évolutifs, productivité végétale.
Ivanishchev Viktor Vasiljevich, docteur en biologie, chercheur principal, directeur de la chaire, [email protégé], Russie, Toula, Université pédagogique de l'État de Toula Lev Tolstoï
Puisque dans le cycle de Calvin, les principaux produits de l’incorporation du carbone inorganique dans le carbone organique sont des composés à trois carbones, ce processus est appelé voie C-3 de la photosynthèse.
Il est important de noter que six révolutions du cycle de Calvin doivent se produire pour synthétiser une molécule de glucose. Chaque chiffre d'affaires utilise trois molécules d'ATP (deux pour activer deux molécules d'acide phosphoglycérique et une pour régénérer le diphosphate de ribulose) et deux molécules de NADP. H 2 pour réduire l'acide en aldéhyde. Ainsi, pour synthétiser une molécule de glucose, il est extrêmement important de dépenser 12 molécules de NADP. Molécules H 2 et 18 ATP.
Il est important de noter que la signification physiologique du cycle de Calvin n'est pas seulement l'acceptation du dioxyde de carbone, mais aussi la création d'une masse de composés glucidiques, qui sont utilisés à la fois pour la synthèse de substances de réserve et pour la création de composants chloroplastiques. et le métabolisme actuel de la cellule. .
La plupart des plantes absorbent le carbone inorganique tout au long du cycle de Calvin. Dans le même temps, un groupe assez important de plantes (environ 500 espèces) d'origine tropicale ont développé au cours de leur évolution des modifications du processus, assimilant du carbone inorganique à travers la formation suite à son acceptation de quatre carbones Connexions. Ce sont des plantes qui se sont adaptées à la photosynthèse dans des conditions de température de l'air élevée et de lumière excessive, ainsi que de faible humidité du sol (sécheresse). Parmi les plantes cultivées, le maïs, le mil, le sorgho et la canne à sucre présentent un tel processus métabolique. Un certain nombre de mauvaises herbes présentent également cette particularité du métabolisme (amarante, millet, glandée), etc.
Une caractéristique de la structure anatomique de ces plantes est la présence de deux types de cellules photosynthétiques, disposées sous la forme de cercles concentriques - les cellules du parenchyme pariétal et du mésophylle, situées radialement autour des faisceaux vasculaires. Ce type de structure anatomique est généralement appelé type Kranz (de l'allemand Kranz - couronne).
Ce type de métabolisme a été étudié dans les années 60 du siècle dernier, avec les recherches des scientifiques soviétiques Karpilov, Nezgovorova, Tarchevsky, ainsi que des scientifiques australiens Hatch et Slack, jouant un rôle majeur. Ce sont eux qui ont proposé un schéma complet du cycle ; à cet égard, il est d'usage d'appeler également ce processus le cycle Hatch-Slack-Karpilov.
Le processus se déroule en deux étapes : le CO 2 entrant dans la mésophylle se combine avec un composé à trois carbones (PEP) - l'acide phosphoénolpyruvique - qui est converti en un composé à quatre carbones. C'est le point clé du processus, d'où son nom, puisque le carbone inorganique, lorsqu'il est accepté par un composé à trois carbones, est converti en un composé à quatre carbones. Compte tenu de la dépendance en laquelle le carbone inorganique composé à quatre carbones est converti, trois groupes de plantes sont distingués :
Le NADP-MDH forme de l'acide malique avec la participation de l'enzyme malate déshydrogénase, puis de l'acide pyruvique,
NAD-MDH forme de l'acide aspartique et de l'alanine,
Le PEP-KK forme de l'acide aspartique et de l'acide phosphoénolpyruvique.
Les plantes les plus importantes pour l'agriculture appartiennent au type NADP-MDG.
Après la formation d'un composé à quatre carbones, celui-ci se déplace dans les cellules internes du parenchyme pariétal et se produit le clivage ou la décarboxylation de cette molécule. Le groupe carboxyle séparé sous forme de COO entre dans le cycle de Calvin et la molécule restante à trois carbones - PEP - retourne à nouveau dans les cellules du mésophylle.
Cette façon de fixer le dioxyde de carbone permet aux plantes d'accumuler des réserves de carbone sous forme d'acides organiques et de réaliser le processus de photosynthèse pendant les heures les plus chaudes de la journée tout en réduisant les pertes d'eau par transpiration dues à la fermeture des stomates. L’efficacité de l’utilisation de l’eau par ces plantes est deux fois plus élevée que celle des plantes originaires des latitudes tempérées.
Les plantes C4 se caractérisent par l'absence de flux inverse de dioxyde de carbone lors de la photorespiration et par un niveau accru de synthèse et d'accumulation de substances organiques.