adaptirane biljke. Glavne faze evolucije biljaka i životinja. Vrste biljaka u odnosu na vodni režim
Pročitajte također
Sada kada smo se upoznali s razlikovnim karakteristikama četiri glavne grupe biljaka, naime briofiti, paprati, golosjemenke i kritosjemenke (cvjetnice), lakše nam je zamisliti evolucijski napredak koji su biljke ostvarile u procesu prilagođavanje životu na kopnu.
Problemi
Možda je najteži problem koji je trebalo nekako prevazići da bi se sa vodenog načina života prešlo na kopneni bio problem dehidracija. Svaka biljka koja nije zaštićena na ovaj ili onaj način, na primjer, nije prekrivena voštanom kutikulom, vrlo brzo će se osušiti i nesumnjivo uginuti. Čak i ako se ova poteškoća prevaziđe, ostali neriješeni problemi ostaju. I prije svega pitanje kako uspješno izvršiti seksualnu reprodukciju. U prvim biljkama u razmnožavanju su učestvovale muške gamete, sposobne da se približe ženskim polnim ćelijama samo plivajući u vodi.
Obično se vjeruje da su prve biljke koje su ovladale zemljom potekle od zelenih algi, kod nekih od evolucijski najnaprednijih predstavnika čiji su se reproduktivni organi pojavili, a to su arhegonija (ženka) i anteridija (muškarci); u ovim organima su polne ćelije bile skrivene i, posljedično, zaštićene. Ova okolnost i niz drugih dobro definiranih uređaja koji pomažu u izbjegavanju isušivanja omogućili su nekim predstavnicima zelenih algi da preuzmu zemljište.
Jedan od najvažnijih evolucijskih trendova biljaka je njihova postupna sve veća neovisnost od vode.
U nastavku su navedene glavne poteškoće povezane s prijelazom iz vodenog u kopneno postojanje.
- Dehidracija. Vazduh je medij za sušenje, a voda je neophodna za život iz raznih razloga (odjeljak 3.1.2). Stoga postoji potreba za uređajima za dobijanje i skladištenje vode.
- Reprodukcija. Osjetljive zametne stanice moraju biti zaštićene, a pokretne muške gamete (spermatozoidi) mogu se susresti sa ženskim spolnim stanicama samo u vodi.
- Podrška. Za razliku od vode, zrak ne može podržavati biljke.
- Ishrana. Biljke trebaju svjetlost i ugljični dioksid (CO 2 ) za fotosintezu, tako da barem dio biljke mora biti iznad zemlje. Međutim, mineralne soli i voda nalaze se u tlu ili na njegovoj površini, a da bi se te tvari efikasno koristile, dio biljke mora biti u zemlji i rasti u mraku.
- Razmjena plina. Za fotosintezu i disanje potrebno je da se razmjena ugljičnog dioksida i kisika odvija ne s okolnim rastvorom, već sa atmosferom.
- faktori životne sredine. Voda, posebno kada je ima toliko, kao, recimo, u jezeru ili okeanu, obezbeđuje visoku postojanost uslova sredine. Kopneno stanište, s druge strane, karakteriše u mnogo većoj meri varijabilnost tako važnih faktora kao što su temperatura, intenzitet svetlosti, koncentracija jona i pH.
Jetrenjak i mahovine
Mahovine su dobro prilagođene širenju spora u kopnenim uslovima: zavisi od sušenja kutije i raspršivanja malih, laganih spora vetrom. Međutim, ove biljke i dalje ovise o vodi iz sljedećih razloga.
- Potrebna im je voda za reprodukciju jer spermatozoidi moraju plivati do arhegonijuma. Ove biljke su razvile adaptacije koje im omogućavaju da oslobađaju spermu samo u vlažnom okruženju, jer se samo u takvom okruženju otvaraju anteridije. Ove biljke su se djelomično prilagodile kopnenom životu, jer su njihove gamete formirane u zaštitnim strukturama - anteridijama i arhegonijama.
- Nemaju posebna potporna tkiva, pa je rast biljke prema gore ograničen.
- Briofiti nemaju korijenje koje može prodrijeti daleko u supstrat, a mogu živjeti samo tamo gdje ima dovoljno vlage i mineralnih soli na površini tla ili u njegovim gornjim slojevima. Međutim, oni imaju rizoide kojima se pričvršćuju za tlo; ovo je jedna od adaptacija na život na čvrstoj podlozi.
2.4. Jetrenjače i mahovine često se nazivaju vodozemcima (vodozemcima) biljnog svijeta. Objasnite ukratko zašto.
paprati
2.5. Paprati su se bolje prilagodile životu na kopnu od jetrenjaka i mahovina. Kako je prikazano?
2.6. Koje su bitne karakteristike mahovina, paprati i jetrenjaka koji su slabo prilagođeni životu na kopnu?
Sjemenice - četinari i cvjetnice
Jedna od glavnih poteškoća s kojima se biljke suočavaju na kopnu je povezana s ranjivosti generacije gametofita. Na primjer, kod paprati, gametofit je osjetljiva izraslina koja proizvodi muške gamete (sperme) kojima je potrebna voda da dođu do jajeta. Međutim, u sjemenskim biljkama gametofit je zaštićen i znatno smanjen.
Sjemenske biljke imaju tri važne prednosti: prvo, heterogene su; drugo, pojava muških spolnih stanica koje ne plivaju i, treće, stvaranje sjemena.
RAZNOLIKOST I MUŠKARSKE GAMETE ZA NEPLIVANJE.
Rice. 2.34. Generalizirana shema životnog ciklusa biljaka, koja odražava smjenu generacija. Obratite pažnju na prisustvo haploidnog (n) i diploidnog (2n) stadijuma. Gametofit je uvijek haploidan i uvijek formira gamete mitotskom diobom. Sporofit je uvijek diploidan i uvijek formira spore kao rezultat mejotičke diobe.
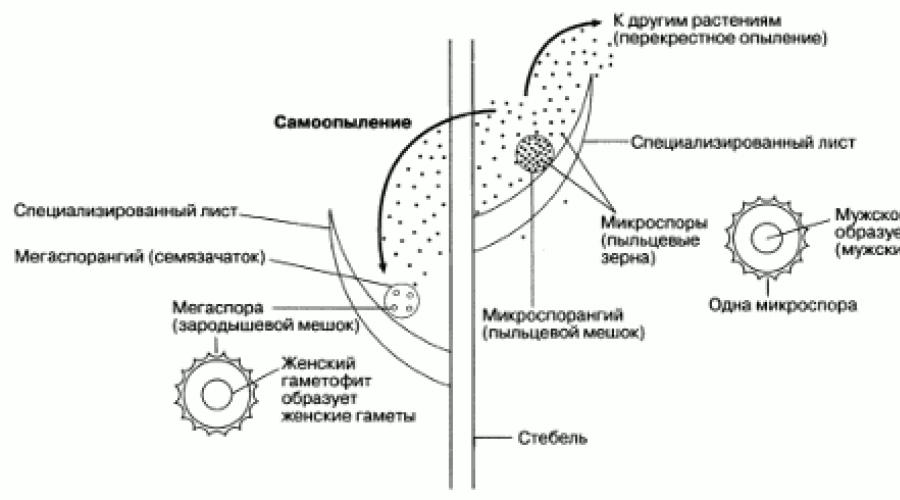
Vrlo važnu ulogu u evoluciji biljaka odigrala je pojava nekih paprati i njihovih bliskih srodnika, koje tvore spore dvije vrste. Ovaj fenomen se zove heterogenost, a biljke su heterosporne. Sve sjemenske biljke su heterosporne. Formiraju velike spore tzv megaspore, u sporangijama jednog tipa (megasporangije) i male spore, zvane mikrospore, u drugom tipu sporangija (mikrosporangije). Klijajući, spore formiraju gametofite (slika 2.34). Megaspore se razvijaju u ženske gametofite, mikrospore u muške. U sjemenskim biljkama, gametofiti formirani od megaspora i mikrospora su vrlo male veličine i nikada se ne oslobađaju od spora. Tako su gametofiti zaštićeni od isušivanja, što je važno evoluciono dostignuće. Međutim, sperma iz muškog gametofita i dalje se mora preseliti u ženski gametofit, što je uvelike olakšano širenjem mikrospora. Budući da su vrlo male, mogu se formirati u velikom broju i vjetar ih prenosi daleko od roditeljskog sporofita. Igrom slučaja, mogu biti u neposrednoj blizini megaspore, koja se u sjemenskim biljkama ne odvaja od roditeljskog sporofita (slika 2.45). To je upravo način na koji se to dešava oprašivanje kod biljaka čija su polenova zrna mikrospore. Muške gamete nastaju u polenovim zrncima.

Rice. 2.45. Šematski prikaz glavnih elemenata diverziteta i oprašivanja.
Biljke sjemena razvile su još jednu evolucijsku prednost. Muške gamete više ne moraju plivati do ženskih spolnih stanica jer su sjemenske biljke razvile polenove cijevi. Razvijaju se iz polenovih zrnaca i rastu prema ženskim gametama. Kroz ovu cijev, muške gamete dospiju do ženske gamete i oplode je. Plutajuća sperma se više ne formira, već su samo muška jezgra uključena u oplodnju.
Shodno tome, biljke su razvile mehanizam oplodnje koji je neovisan o vodi. To je bio jedan od razloga zašto su sjemenske biljke bile toliko superiorne u odnosu na druge biljke u razvoju zemljišta. U početku se oprašivanje odvijalo samo uz pomoć vjetra - prilično nasumičan proces, praćen velikim gubicima polena. Međutim, već u ranim fazama evolucije, prije oko 300 miliona godina u periodu karbona, pojavili su se leteći insekti, a sa njima i mogućnost efikasnijeg oprašivanja. Cvjetnice uveliko koriste oprašivanje insektima, dok oprašivanje vjetrom još uvijek prevladava kod četinara.
SJEME. U ranim heterosporoznim biljkama, megaspore su se oslobađale iz roditeljskog sporofita poput mikrospora. Kod sjemenskih biljaka megaspore se ne odvajaju od matične biljke, ostaju u megasporangiji ili ovules(Sl. 2.45). Ovala sadrži žensku gametu. Nakon oplodnje ženske gamete, ovule se već nazivaju sjeme. Dakle, sjeme je oplođena ovula. Prisutnost ovule i sjemena daje određene prednosti sjemenskim biljkama.
- Ženski gametofit zaštićen je ovulom. Potpuno je ovisan o roditeljskom sporofitu i, za razliku od slobodnoživućeg gametofita, neosjetljiv je na dehidraciju.
- Nakon oplodnje, sjeme formira rezervu hranjivih tvari koju gametofit prima od matične biljke sporofita, od koje se još uvijek ne odvaja. Ovu rezervu koristi zigota u razvoju (sljedeća generacija sporofita) nakon klijanja sjemena.
- Seme je dizajnirano da preživi nepovoljne uslove i ostane u stanju mirovanja sve dok uslovi ne budu povoljni za klijanje.
- Sjeme može razviti različite adaptacije kako bi se olakšalo njihovo širenje.
Sjeme je složena struktura u kojoj su sastavljene ćelije tri generacije - roditeljski sporofit, ženski gametofit i embrion sljedeće generacije sporofita. Roditeljski sporofit obezbjeđuje sjemenu sve što mu je potrebno za život, i to tek nakon što sjeme potpuno sazre, tj. akumulira zalihe hranljivih materija za embrion sporofita, odvaja se od roditeljskog sporofita.
2.7. Šanse za opstanak i razvoj polenovih zrnaca (mikrospora) koje se prenose vjetrom su mnogo manje nego za spore Dryopteris. Zašto?
2.8. Objasni zašto su megaspore velike, a mikrospore male.
2.7.7. Kratak popis prilagođavanja sjemenskih biljaka životu na kopnu
Glavne prednosti sjemenskih biljaka u odnosu na sve ostale su sljedeće.
- Generacija gametofita je znatno smanjena i potpuno ovisi o sporofitu, dobro prilagođenom životu na kopnu, unutar kojeg je gametofit uvijek zaštićen. Kod drugih biljaka gametofit se vrlo lako suši.
- Oplodnja se odvija nezavisno od vode. Muške gamete su nepokretne i raspršene unutar polenovih zrna vjetrom ili insektima. Konačni prijenos muških spolnih stanica na ženske spolne stanice događa se uz pomoć polenove cijevi.
- Oplođene ovule (sjemenke) ostaju neko vrijeme na roditeljskom sporofitu, od kojeg dobijaju zaštitu i hranu prije nego što se rasprše.
- Kod mnogih sjemenskih biljaka uočava se sekundarni rast sa taloženjem velikih količina drveta koje ima potpornu funkciju. Takve biljke izrastaju u drveće i grmlje koje se može efikasno nadmetati za svjetlo i druge resurse.
Neki od najvažnijih evolucijskih trendova sumirani su na sl. 2.33. Biljke sjemena imaju i druge osobine koje su svojstvene biljkama ne samo ove grupe, već igraju i ulogu prilagodbe životu na kopnu.

Rice. 2.33. Sistematika biljaka i neki glavni trendovi u evoluciji biljaka.
- Pravi korijeni osiguravaju izvlačenje vlage iz tla.
- Biljke su od isušivanja zaštićene epidermom sa vodonepropusnom kutikulom (ili čepom koji se formira nakon sekundarnog rasta).
- Epidermu kopnenih dijelova biljke, posebno listova, probijaju mnoge sitne proreze tzv. stomata kroz koje se odvija razmjena gasova između postrojenja i atmosfere.
- Biljke takođe imaju specijalizovane adaptacije na život u vrućim sušnim uslovima (pogl. 19 i 20).
Prilagodljivost ontogeneze biljaka na uslove sredine rezultat je njihovog evolucionog razvoja (varijabilnost, nasljednost, selekcija). Tokom filogeneze svake biljne vrste, u procesu evolucije, razvile su se određene potrebe pojedinca za uslovima postojanja i prilagodljivosti ekološkoj niši koju zauzima. Tolerantnost na vlagu i hlad, otpornost na toplotu, hladnoću i druge ekološke karakteristike pojedinih biljnih vrsta formirane su tokom evolucije kao rezultat dugotrajnog izlaganja odgovarajućim uslovima. Dakle, biljke koje vole toplinu i biljke kratkog dana karakteristične su za južne geografske širine, manje zahtjevne za toplinu i biljke dugog dana - za sjeverne.
U prirodi, u jednoj geografskoj regiji, svaka biljna vrsta zauzima ekološku nišu koja odgovara njenim biološkim karakteristikama: voli vlagu - bliže vodenim tijelima, tolerantna na sjenu - pod krošnjom šume, itd. Nasljednost biljaka se formira pod utjecajem određenih uslova životne sredine. Važni su i vanjski uslovi ontogeneze biljaka.
U većini slučajeva, biljke i usjevi (zasadi) poljoprivrednih kultura, doživljavajući djelovanje određenih nepovoljnih faktora, pokazuju otpor prema njima kao rezultat prilagođavanja uvjetima postojanja koji su se povijesno razvijali, što je primijetio K. A. Timiryazev.
1. Osnovne životne sredine.
Prilikom proučavanja životne sredine (staništa biljaka i životinja i ljudskih proizvodnih aktivnosti) izdvajaju se sledeće glavne komponente: vazdušna sredina; vodena sredina (hidrosfera); fauna (ljudi, domaće i divlje životinje, uključujući ribe i ptice); flora (kultivisane i samonikle biljke, uključujući i one koje rastu u vodi), tlo (vegetacijski sloj), podzemlje (gornji dio zemljine kore, unutar kojeg je moguće rudarenje); klimatsko i akustičko okruženje.
Vazdušna sredina može biti eksterna, u kojoj većina ljudi provodi manji dio svog vremena (do 10-15%), interna proizvodna (u njoj osoba provodi do 25-30% vremena) i unutrašnja stambena, gdje ljudi ostaju većinu vremena (do 60 -70% ili više).
Vanjski vazduh na površini zemlje sadrži po zapremini: 78,08% azota; 20,95% kiseonika; 0,94% inertnih plinova i 0,03% ugljičnog dioksida. Na visini od 5 km sadržaj kiseonika ostaje isti, dok se azota povećava na 78,89%. Često zrak u blizini površine zemlje ima razne nečistoće, posebno u gradovima: tamo sadrži više od 40 sastojaka koji su stranci prirodnom zračnom okruženju. Unutrašnji vazduh u stanovima, po pravilu, ima
povećan sadržaj ugljičnog dioksida, a unutrašnji zrak industrijskih prostorija obično sadrži nečistoće, čija je priroda određena tehnologijom proizvodnje. Među plinovima se oslobađa vodena para, koja ulazi u atmosferu kao rezultat isparavanja sa Zemlje. Najveći dio (90%) koncentrisan je u najnižem pet-kilometarskom sloju atmosfere, s visinom se njegova količina vrlo brzo smanjuje. Atmosfera sadrži mnogo prašine koja tamo stiže sa površine Zemlje i dijelom iz svemira. Za vrijeme jakih valova vjetrovi podižu vodeni mlaz iz mora i okeana. Tako čestice soli iz vode dospiju u atmosferu. Kao rezultat vulkanskih erupcija, šumskih požara, industrijskih objekata itd. vazduh je zagađen produktima nepotpunog sagorevanja. Najviše prašine i drugih nečistoća nalazi se u prizemnom sloju vazduha. Čak i nakon kiše, 1 cm sadrži oko 30 hiljada čestica prašine, a po suhom ih je nekoliko puta više po suhom vremenu.
Sve ove sitne nečistoće utiču na boju neba. Molekuli gasova raspršuju kratkotalasni dio spektra sunčevog zraka, tj. ljubičastih i plavih zraka. Dakle, tokom dana je nebo plavo. A čestice nečistoća, koje su mnogo veće od molekula plina, raspršuju svjetlosne zrake gotovo svih valnih dužina. Stoga, kada je zrak prašnjav ili sadrži kapljice vode, nebo postaje bjelkasto. Na velikim visinama nebo je tamnoljubičasto, pa čak i crno.
Kao rezultat fotosinteze koja se odvija na Zemlji, vegetacija godišnje formira 100 milijardi tona organskih supstanci (oko polovine otpada na mora i okeane), dok asimilira oko 200 milijardi tona ugljičnog dioksida i ispušta oko 145 milijardi tona u okruženje. slobodnog kiseonika, veruje se da fotosintezom nastaje sav kiseonik u atmosferi. O ulozi zelenih površina u ovom ciklusu govore sledeći podaci: 1 hektar zelenih površina čisti vazduh od 8 kg ugljen-dioksida u proseku za 1 sat (200 ljudi emituje za to vreme prilikom disanja). Odraslo drvo oslobađa 180 litara kiseonika dnevno, a za pet meseci (od maja do septembra) apsorbuje oko 44 kg ugljen-dioksida.
Količina oslobođenog kisika i apsorbiranog ugljičnog dioksida ovisi o starosti zelenih površina, sastavu vrsta, gustoći sadnje i drugim faktorima.
Jednako su važne i morske biljke – fitoplankton (uglavnom alge i bakterije), koje fotosintezom oslobađaju kisik.
Vodeni okoliš uključuje površinske i podzemne vode. Površinske vode su uglavnom koncentrisane u okeanu, sa sadržajem od 1 milijardu 375 miliona kubnih kilometara - oko 98% sve vode na Zemlji. Površina okeana (vodno područje) je 361 milion kvadratnih kilometara. To je oko 2,4 puta više od površine zemlje - teritorija koja zauzima 149 miliona kvadratnih kilometara. Voda u okeanu je slana, a najveći dio (više od 1 milijarde kubnih kilometara) zadržava konstantan salinitet od oko 3,5% i temperaturu od oko 3,7°C. Primjetne razlike u salinitetu i temperaturi uočavaju se gotovo isključivo na površini sloju vode, kao iu rubnim i posebno u Sredozemnom moru. Sadržaj otopljenog kisika u vodi značajno opada na dubini od 50-60 metara.
Podzemne vode mogu biti slane, bočate (nižeg saliniteta) i svježe; postojeće geotermalne vode imaju povišenu temperaturu (više od 30ºC).
Za proizvodne aktivnosti čovječanstva i potrebe njegovih domaćinstava potrebna je slatka voda, čija količina iznosi samo 2,7% ukupne količine vode na Zemlji, a njen vrlo mali udio (samo 0,36%) je dostupan na mjestima koja su lako dostupni za vađenje. Većina slatke vode nalazi se u snijegu i slatkovodnim santima leda koji se nalaze u područjima prvenstveno u Antarktičkom krugu.
Godišnji svjetski riječni otjecanje slatke vode iznosi 37,3 hiljade kubnih kilometara. Osim toga, može se koristiti i dio podzemne vode od 13 hiljada kubnih kilometara. Nažalost, najveći dio riječnog toka u Rusiji, koji iznosi oko 5.000 kubnih kilometara, pada na rubne i slabo naseljene sjeverne teritorije.
Klimatsko okruženje je važan faktor koji determiniše razvoj različitih vrsta flore i faune i njenu plodnost. Karakteristična karakteristika Rusije je da većina njene teritorije ima mnogo hladniju klimu nego u drugim zemljama.
Sve razmatrane komponente životne sredine su uključene u
BIOSFERA: Zemljina školjka, uključujući dio atmosfere, hidrosferu i gornji dio litosfere, koji su međusobno povezani složenim biohemijskim ciklusima migracije materije i energije, geološka ljuska Zemlje, naseljena živim organizmima. Gornja granica života biosfere ograničena je intenzivnom koncentracijom ultraljubičastih zraka; niža - visoka temperatura unutrašnjosti zemlje (preko 100`C). Njegove ekstremne granice dostižu samo niži organizmi - bakterije.
Adaptacija (prilagođavanje) biljke specifičnim uslovima sredine obezbeđuje se fiziološkim mehanizmima (fiziološka adaptacija), au populaciji organizama (vrste) - mehanizmima genetske varijabilnosti, nasleđa i selekcije (genetska adaptacija). Faktori okoline se mogu mijenjati redovno i nasumično. Redovno promjenjivi uvjeti okoline (promjena godišnjih doba) razvijaju kod biljaka genetsku adaptaciju na ove uslove.
U prirodnim uslovima rasta ili uzgoja neke vrste, u toku svog rasta i razvoja, često doživljavaju uticaj nepovoljnih faktora sredine, među kojima su temperaturne fluktuacije, suša, prekomerna vlaga, zaslanjenost zemljišta i dr. sposobnost prilagođavanja promenljivim uslovima.uslovi sredine u granicama koje određuje njen genotip. Što je veća sposobnost biljke da mijenja metabolizam u skladu sa okruženjem, to je veća brzina reakcije ove biljke i bolja je sposobnost prilagođavanja. Ovo svojstvo razlikuje otporne sorte poljoprivrednih kultura. Blage i kratkotrajne promjene faktora sredine u pravilu ne dovode do značajnijih poremećaja fizioloških funkcija biljaka, što je posljedica njihove sposobnosti da održavaju relativno stabilno stanje u promjenjivim uvjetima okoline, odnosno održavaju homeostazu. Međutim, oštri i dugotrajni udari dovode do poremećaja mnogih funkcija biljke, a često i do njene smrti.
Pod uticajem nepovoljnih uslova, smanjenje fizioloških procesa i funkcija može dostići kritične nivoe koji ne obezbeđuju realizaciju genetskog programa ontogeneze, narušava se energetski metabolizam, regulacioni sistemi, metabolizam proteina i druge vitalne funkcije biljnog organizma. Kada je biljka izložena nepovoljnim faktorima (stresorima), u njoj nastaje stresno stanje, odstupanje od norme - stres. Stres je opća nespecifična adaptivna reakcija organizma na djelovanje bilo kojeg štetnog faktora. Postoje tri glavne grupe faktora koji izazivaju stres kod biljaka: fizički - nedovoljna ili prekomjerna vlažnost, svjetlost, temperatura, radioaktivno zračenje, mehanički stres; hemikalije - soli, gasovi, ksenobiotici (herbicidi, insekticidi, fungicidi, industrijski otpad, itd.); biološki - oštećenja od patogena ili štetočina, konkurencija s drugim biljkama, utjecaj životinja, cvjetanje, sazrijevanje plodova.
Jačina stresa zavisi od brzine razvoja nepovoljne situacije za biljku i nivoa faktora stresa. Sa sporim razvojem nepovoljnih uslova, biljka se bolje prilagođava njima nego kratkotrajnim, ali snažnim dejstvom. U prvom slučaju, u pravilu se u većoj mjeri manifestiraju specifični mehanizmi rezistencije, u drugom - nespecifični.
U nepovoljnim prirodnim uslovima otpornost i produktivnost biljaka određuju se nizom znakova, svojstava i zaštitnih i adaptivnih reakcija. Različite biljne vrste obezbeđuju stabilnost i opstanak u nepovoljnim uslovima na tri glavna načina: kroz mehanizme koji im omogućavaju da izbegnu štetne efekte (mirovanje, efemera, itd.); kroz posebne konstrukcijske uređaje; zbog fizioloških svojstava koja im omogućavaju da savladaju štetne uticaje okoline.
Jednogodišnje poljoprivredne biljke u umjerenim zonama, zaokružujući svoju ontogenezu u relativno povoljnim uslovima, prezimljuju u obliku stabilnog sjemena (dormant). Mnoge višegodišnje biljke prezimljuju kao podzemni skladišni organi (lukovice ili rizomi) zaštićeni od smrzavanja slojem zemlje i snijega. Voćke i grmlje umjerenih zona, štiteći se od zimske hladnoće, osipaju lišće.
Zaštita od nepovoljnih faktora okoline u biljkama je obezbeđena strukturnim adaptacijama, osobinama anatomske strukture (kutikula, kora, mehanička tkiva i dr.), posebnim zaštitnim organima (sagorevanje dlaka, bodlji), motoričkim i fiziološkim reakcijama i proizvodnjom zaštitnih supstance (smole, fitoncidi, toksini, zaštitni proteini).
Strukturne adaptacije uključuju sitno lišće, pa čak i odsustvo listova, voštanu kutikulu na površini listova, njihovo gusto izostavljanje i uranjanje puči, prisustvo sočnih listova i stabljika koji zadržavaju rezerve vode, erektoidno ili viseće listove itd. imaju različite fiziološke mehanizme koji im omogućavaju da se prilagode nepovoljnim uslovima.uslovi sredine. Ovo je samo-tip fotosinteze u sukulentnim biljkama, koji minimizira gubitak vode i neophodan je za opstanak biljaka u pustinji, itd.
2. Adaptacija u biljkama
Tolerantnost biljaka na hladnoću
Otpornost biljaka na niske temperature dijeli se na otpornost na hladnoću i otpornost na mraz. Pod otpornošću na hladnoću podrazumijeva se sposobnost biljaka da podnose pozitivne temperature nešto veće od 0 C. Hladnootpornost je karakteristična za biljke umjerenog pojasa (ječam, zob, lan, grahorica i dr.). Tropske i suptropske biljke oštećuju se i umiru na temperaturama od 0º do 10º C (kafa, pamuk, krastavac itd.). Za većinu poljoprivrednih biljaka niske pozitivne temperature nisu štetne. To je zbog činjenice da se tijekom hlađenja enzimski aparat biljaka ne poremeti, otpornost na gljivične bolesti se ne smanjuje i uopće ne dolazi do primjetnih oštećenja biljaka.
Stepen otpornosti na hladnoću različitih biljaka nije isti. Mnoge biljke južnih geografskih širina su oštećene hladnoćom. Na temperaturi od 3°C oštećuju se krastavac, pamuk, pasulj, kukuruz, patlidžan. Sorte se razlikuju po otpornosti na hladnoću. Za karakterizaciju hladno otpornosti biljaka koristi se koncept temperaturnog minimuma na kojem prestaje rast biljaka. Za veliku grupu poljoprivrednih biljaka njegova vrijednost je 4 °C. Međutim, mnoge biljke imaju viši temperaturni minimum i stoga su manje otporne na hladnoću.
Adaptacija biljaka na niske pozitivne temperature.
Otpornost na niske temperature je genetski određena osobina. Otpornost biljaka na hladnoću određena je sposobnošću biljaka da održe normalnu strukturu citoplazme, da mijenjaju metabolizam tokom perioda hlađenja i naknadnog povećanja temperature na dovoljno visokom nivou.
Otpornost biljaka na mraz
Otpornost na mraz - sposobnost biljaka da tolerišu temperature ispod 0 ° C, niske negativne temperature. Biljke otporne na mraz su u stanju spriječiti ili smanjiti učinak niskih negativnih temperatura. Mrazevi zimi sa temperaturama ispod -20°C uobičajeni su za značajan dio teritorije Rusije. Jednogodišnje, dvogodišnje i višegodišnje biljke su izložene mrazu. Biljke podnose zimske uslove u različitim periodima ontogeneze. U jednogodišnjim usjevima prezimljuju sjemenke (jare), klijalice (ozime), u dvogodišnjim i višegodišnjim usjevima - gomolji, korjenasti usjevi, lukovice, rizomi, odrasle biljke. Sposobnost prezimljavanja ozimih, višegodišnjih zeljastih i drvenastih voćaka je zbog njihove prilično visoke otpornosti na mraz. Tkiva ovih biljaka mogu se smrznuti, ali biljke ne umiru.
Zamrzavanje biljnih ćelija i tkiva i procesi koji se pri tome dešavaju.
Sposobnost biljaka da podnose negativne temperature određena je nasljednom osnovom određene biljne vrste, međutim otpornost na mraz jedne te iste biljke ovisi o uvjetima koji prethode nastanku mraza, što utječe na prirodu stvaranja leda. Led se može formirati i u protoplastu ćelije i u međućelijskom prostoru. Ne uzrokuje svako stvaranje leda odumiranje biljnih stanica.
Postepeno smanjenje temperature brzinom od 0,5-1 °C/h dovodi do stvaranja kristala leda, prvenstveno u međućelijskim prostorima, koji u početku ne izazivaju smrt ćelije. Međutim, posljedice ovog procesa mogu biti štetne za ćeliju. Formiranje leda u protoplastu ćelije, u pravilu, nastaje brzim smanjenjem temperature. Dolazi do koagulacije protoplazmatskih proteina, ćelijske strukture su oštećene kristalima leda koji se formiraju u citosolu, ćelije umiru. Biljke ubijene mrazom nakon odmrzavanja gube turgor, voda istječe iz njihovih mesnatih tkiva.
Biljke otporne na mraz imaju adaptacije koje smanjuju dehidraciju stanica. Sa smanjenjem temperature u takvim biljkama, primjećuje se povećanje sadržaja šećera i drugih tvari koje štite tkiva (krioprotektori), prvenstveno hidrofilni proteini, mono- i oligosaharidi; smanjenje hidratacije ćelija; povećanje količine polarnih lipida i smanjenje zasićenosti njihovih ostataka masnih kiselina; povećanje broja zaštitnih proteina.
Na stepen otpornosti biljaka na mraz u velikoj meri utiču šećeri, regulatori rasta i druge supstance koje se formiraju u ćelijama. Kod biljaka koje prezimljuju, šećeri se nakupljaju u citoplazmi, a sadržaj škroba se smanjuje. Utjecaj šećera na povećanje otpornosti biljaka na mraz je višestruk. Akumulacija šećera sprečava smrzavanje velike količine intracelularne vode, značajno smanjuje količinu leda koji se formira.
Svojstvo otpornosti na mraz formira se u procesu ontogeneze biljke pod utjecajem određenih uvjeta okoline u skladu s genotipom biljke, povezano s naglim smanjenjem stope rasta, prijelazom biljke u stanje mirovanja.
Životni ciklus razvoja zimskih, dvogodišnjih i višegodišnjih biljaka kontroliran je sezonskim ritmom svjetlosnih i temperaturnih perioda. Za razliku od proljetnih jednogodišnjih biljaka, one se počinju pripremati da izdrže nepovoljne zimske uslove od trenutka kada prestanu rasti, a zatim tokom jeseni kada temperature padnu.
Zimska otpornost biljaka
Zimljivost kao otpornost na kompleks nepovoljnih faktora prezimljavanja.
Direktno djelovanje mraza na ćelije nije jedina opasnost koja prijeti višegodišnjim zeljastim i drvenastim usjevima, ozimim biljkama tokom zime. Pored direktnog dejstva mraza, biljke su izložene i nizu drugih nepovoljnih faktora. Temperature mogu značajno da variraju tokom zime. Mrazevi se često zamjenjuju kratkotrajnim i dugotrajnim otapanjem. Zimi snježne oluje nisu neuobičajene, a u zimama bez snijega u južnijim krajevima zemlje javljaju se i suvi vjetrovi. Sve to iscrpljuje biljke koje nakon prezimljavanja izlaze vrlo oslabljene i mogu naknadno uginuti.
Posebno brojne štetne efekte imaju zeljaste višegodišnje i jednogodišnje biljke. Na teritoriji Rusije, u nepovoljnim godinama, smrt ozimih žitarica dostiže 30-60%. Ne umiru samo ozimi usevi, već i višegodišnje trave, plantaže voća i jagodičastog voća. Osim niskih temperatura, zimske biljke oštećuju i umiru od niza drugih nepovoljnih faktora zimi i u rano proljeće: vlaženje, vlaženje, ledena kora, ispupčenje, oštećenja od zimske suše.
Vlaženje, natapanje, smrt pod ledenom korom, ispupčenje, oštećenja od zimske suše.
Damping out. Među navedenim nedaćama, prvo mjesto zauzima propadanje biljaka. Smrt biljaka od prigušivanja uočava se uglavnom u toplim zimama sa velikim snježnim pokrivačem koji traje 2-3 mjeseca, posebno ako snijeg pada na vlažno i odmrznuto tlo. Istraživanja su pokazala da je uzrok uginuća ozimih usjeva od prigušenja, iscrpljivanje biljaka. Nalazeći se pod snijegom na temperaturi od oko 0°C u visoko vlažnom okruženju, gotovo potpunom mraku, odnosno u uslovima u kojima je proces disanja prilično intenzivan i fotosinteza isključena, biljke postepeno troše šećer i druge rezerve hranljivih materija nakupljene tokom perioda. prolazeći kroz prvu fazu stvrdnjavanja, umiru od iscrpljenosti (sadržaj šećera u tkivima se smanjuje sa 20 na 2-4%) i proljetnih mrazeva. Takve biljke lako oštete snježna plijesan u proljeće, što također dovodi do njihove smrti.
Vlaženje. Vlaženje se javlja uglavnom u proljeće na niskim mjestima u periodu otapanja snijega, rjeđe tokom dugotrajnog odmrzavanja, kada se na površini tla nakuplja otopljena voda, koja se ne upija u smrznuto tlo i može poplaviti biljke. U ovom slučaju uzrok smrti biljke je oštar nedostatak kisika (anaerobni uvjeti - hipoksija). Kod biljaka koje se nalaze ispod sloja vode normalno disanje prestaje zbog nedostatka kiseonika u vodi i zemljištu. Nedostatak kisika pojačava anaerobno disanje biljaka, uslijed čega se mogu stvarati otrovne tvari i biljke umiru od iscrpljenosti i direktnog trovanja organizma.
Smrt pod ledenom korom. Ledena kora se formira na poljima u područjima gdje se česta odmrzavanja zamjenjuju jakim mrazevima. Učinak namakanja u ovom slučaju može biti pogoršan. U tom slučaju dolazi do stvaranja visećih ili mljevenih (kontaktnih) ledenih kora. Viseće kore su manje opasne, jer se formiraju na vrhu tla i praktički ne dolaze u kontakt s biljkama; lako ih je uništiti valjkom.
Kada se formira kontinuirana ledena kontaktna kora, biljke se potpuno smrzavaju u led, što dovodi do njihove smrti, jer su biljke, već oslabljene od natapanja, podvrgnute vrlo jakom mehaničkom pritisku.
Ispupčenje. Oštećenje i smrt biljaka od ispupčenja određuju se rupturama u korijenskom sistemu. Izbočenje biljaka se uočava ako se mrazevi javljaju u jesen u nedostatku snježnog pokrivača ili ako ima malo vode u površinskom sloju tla (tokom jesenje suše), kao i tokom odmrzavanja, ako snježna voda ima vremena da se upije u tlo. U tim slučajevima smrzavanje vode ne počinje s površine tla, već na određenoj dubini (gdje ima vlage). Sloj leda koji se formira na dubini postepeno se zgušnjava zbog kontinuiranog protoka vode kroz kapilare tla i podiže (ispupčuje) gornje slojeve tla zajedno sa biljkama, što dovodi do lomljenja korijena biljaka koje imaju prodro do znatne dubine.
Šteta od zimske suše. Stabilan snježni pokrivač štiti ozime žitarice od isušivanja zimi. Međutim, u uslovima bezsnežne ili malo snežne zime, poput voćaka i žbunja, u nizu regiona Rusije često su u opasnosti od prekomernog isušivanja stalnim i jakim vetrovima, posebno krajem zime sa značajnim zagrevanjem sunce. Činjenica je da se vodni bilans biljaka zimi razvija izuzetno nepovoljno, jer protok vode iz smrznutog tla praktički prestaje.
Da bi se smanjilo isparavanje vode i štetni efekti zimske suše, vrste voćaka formiraju debeli sloj plute na granama i osipaju lišće za zimu.
Vernalizacija
Fotoperiodični odgovori na sezonske promjene dužine dana važni su za učestalost cvjetanja mnogih vrsta u umjerenim i tropskim regijama. Međutim, treba napomenuti da je među vrstama umjerenih geografskih širina koje pokazuju fotoperiodične reakcije relativno malo onih koje cvjetaju u proljeće, iako se stalno susrećemo sa značajnim brojem „cvijeća koje cvjeta u proljeće“, a mnogi od ovih proljetnocvjetnih oblika , na primjer, Ficariaverna, jaglac (Primulavutgaris), ljubičice (vrste roda Viola) i dr. pokazuju izraženo sezonsko ponašanje, ostajući vegetativno do kraja godine nakon obilnog proljetnog cvjetanja. Može se pretpostaviti da je proljetno cvjetanje reakcija na kratke dane zimi, ali za mnoge vrste to izgleda nije slučaj.
Naravno, dužina dana nije jedini vanjski faktor koji se mijenja tokom godine. Jasno je da temperatura takođe pokazuje izražene sezonske varijacije, posebno u umjerenim područjima, iako ovaj faktor pokazuje značajne fluktuacije, kako dnevne tako i godišnje. Znamo da sezonske promjene temperature, kao i promjene dužine dana, imaju značajan utjecaj na cvjetanje mnogih biljnih vrsta.
Vrste biljaka kojima je potrebno hlađenje da bi prešle na cvjetanje.
Utvrđeno je da mnoge vrste, uključujući zimske jednogodišnje biljke, kao i dvogodišnje i višegodišnje zeljaste biljke, moraju biti rashlađene da bi prešle na cvjetanje.
Poznato je da su zimske i dvogodišnje biljke monokarpne biljke koje zahtijevaju jarovizaciju - one ostaju vegetativne tokom prve vegetacijske sezone i cvjetaju sljedećeg proljeća ili ranog ljeta kao odgovor na period hlađenja primljen zimi. Potreba za hlađenjem dvogodišnjih biljaka za izazivanje cvjetanja eksperimentalno je dokazana kod brojnih vrsta kao što su cvekla (Betavulgaris), celer (Apiutngraveolens), kupus i druge kultivisane sorte iz roda Brassica, zvončić (Campanulamedium), mjesečeva trava (Lunariabiennis) , lisičarka (Digitalispurpurea) i dr. Ako se biljke digitalisa, koje se u normalnim uvjetima ponašaju kao dvogodišnje, odnosno cvjetaju u drugoj godini nakon nicanja, drže u stakleniku, mogu ostati vegetativne nekoliko godina. U područjima sa blagim zimama, kelj može rasti na otvorenom nekoliko godina bez "vrh strijele" (tj. cvjetanja) u proljeće, što se obično događa u područjima sa hladnim zimama. Takve vrste nužno zahtijevaju jarovizaciju, ali kod niza drugih vrsta cvjetanje se ubrzava kada su izložene hladnoći, ali može nastati i bez jarovizacije; takve vrste koje pokazuju fakultativnu potrebu za hladnoćom uključuju zelenu salatu (Lactucasaiiva), spanać (Spinacia oleracea) i kasnocvjetajući grašak (Pistimsa-tivum).
Kao i dvogodišnje biljke, mnoge trajnice zahtijevaju izlaganje hladnoći i neće cvjetati bez godišnje zimske hladnoće. Od uobičajenih višegodišnjih biljaka, jaglac (Primulavulgaris), ljubičice (Violaspp.), lacfiol (Cheiranthuscheirii i C. allionii), levka (Mathiolaincarna), neke sorte krizantema (Chrisanthemummorifolium), vrsta turskog carnausstera (D.) , pljeve (Loliumperenne). Višegodišnje vrste zahtijevaju revernalizaciju svake zime.
Vjerovatno je da se može naći i drugim trajnicama koje cvjetaju u proljeće treba hlađenje. Lukovičaste biljke koje cvjetaju u proljeće kao što su narcise, zumbuli, borovnice (Endymionnonscriptus), krokusi, itd. ne zahtijevaju hlađenje do početka cvjetanja jer je prethodno ljeto u lukovicama uspostavljena cvjetna primordija, ali njihov rast u velikoj mjeri ovisi o temperaturnim uvjetima . Na primjer, kod tulipana početku cvjetanja pogoduju relativno visoke temperature (20°C), ali za izduživanje stabljike i rast listova optimalna temperatura u početku je 8-9°C, s postepenim povećanjem u kasnijim fazama. do 13, 17 i 23°C. Slične reakcije na temperaturu karakteristične su za zumbule i narcise.
Kod mnogih vrsta inicijacija cvjetanja se ne odvija tokom samog perioda hlađenja, i počinje tek nakon što je biljka bila izložena višim temperaturama nakon hlađenja.
Dakle, iako se metabolizam većine biljaka znatno usporava na niskim temperaturama, nema sumnje da jarovizacija uključuje aktivne fiziološke procese, čija je priroda još uvijek potpuno nepoznata.
Otpornost biljaka na toplinu
Otpornost na toplinu (tolerantnost na toplinu) - sposobnost biljaka da izdrže djelovanje visokih temperatura, pregrijavanja. Ovo je genetski određena osobina. Biljne vrste se razlikuju po svojoj toleranciji na visoke temperature.
Prema otpornosti na toplinu razlikuju se tri grupe biljaka.
Otporne na toplotu - termofilne modrozelene alge i bakterije toplih mineralnih izvora, sposobne da izdrže temperature do 75-100 °C. Otpornost na toplotu termofilnih mikroorganizama određena je visokim nivoom metabolizma, povećanim sadržajem RNK u ćelijama i otpornošću citoplazmatskog proteina na termičku koagulaciju.
Toplotolerantne - biljke pustinjskih i suhih staništa (sukulenti, neki kaktusi, članovi porodice Crassula), podnose zagrijavanje sunčevom svjetlošću do 50-65ºS. Otpornost sukulenata na toplinu je u velikoj mjeri određena povećanim viskozitetom citoplazme i sadržajem vezane vode u stanicama, te smanjenim metabolizmom.
Neotporne na toplinu - mezofitne i vodene biljke. Mezofiti otvorenih mjesta tolerišu kratkotrajno izlaganje temperaturama od 40-47 °C, zasjenjena mjesta - oko 40-42 °C, vodene biljke podnose temperature do 38-42 °C. Od poljoprivrednih kultura, toplotoljubive biljke južnih geografskih širina (sirak, pirinač, pamuk, ricinus, itd.) su najotpornije na toplotu.
Mnogi mezofiti podnose visoke temperature zraka i izbjegavaju pregrijavanje zbog intenzivne transpiracije koja smanjuje temperaturu listova. Mezofiti otporniji na toplinu odlikuju se povećanom viskoznošću citoplazme i povećanom sintezom proteina enzima otpornih na toplinu.
Biljke su razvile sistem morfoloških i fizioloških adaptacija koje ih štite od termičkih oštećenja: svijetla boja površine koja odražava insolaciju; savijanje i uvijanje listova; pubescencija ili ljuske koje štite dublja tkiva od pregrijavanja; tanki slojevi plutenog tkiva koji štite floem i kambijum; veća debljina kutikularnog sloja; visok sadržaj ugljikohidrata i nizak - vode u citoplazmi itd.
Biljke vrlo brzo reagiraju na toplotni stres induktivnom adaptacijom. Mogu se pripremiti za izlaganje visokim temperaturama za nekoliko sati. Dakle, u toplim danima otpornost biljaka na visoke temperature u popodnevnim satima veća je nego u jutarnjim satima. Obično je ovaj otpor privremen, ne konsoliduje se i nestaje prilično brzo ako se ohladi. Reverzibilnost toplotnog izlaganja može biti u rasponu od nekoliko sati do 20 dana. Tokom formiranja generativnih organa smanjuje se toplotna otpornost jednogodišnjih i dvogodišnjih biljaka.
Otpornost biljaka na sušu
Suše su postale uobičajena pojava u mnogim regionima Rusije i zemalja ZND. Suša je dug period bez kiše, praćen smanjenjem relativne vlažnosti zraka, vlage u tlu i porastom temperature, kada nisu zadovoljene normalne potrebe biljaka za vodom. Na teritoriji Rusije postoje regije nestabilne vlage sa godišnjom količinom padavina od 250-500 mm i sušne regije sa manje od 250 mm padavina godišnje sa stopom isparavanja većom od 1000 mm.
Otpornost na sušu - sposobnost biljaka da izdrže duge sušne periode, značajan deficit vode, dehidraciju ćelija, tkiva i organa. Istovremeno, šteta na usjevu zavisi od trajanja suše i njenog intenziteta. Razlikovati sušu u tlu i atmosfersku sušu.
Suša u tlu je uzrokovana dugotrajnim nedostatkom kiše u kombinaciji s visokom temperaturom zraka i sunčevom insolacijom, pojačanim isparavanjem s površine tla i transpiracijom te jakim vjetrovima. Sve to dovodi do isušivanja korijenskog sloja tla, smanjenja opskrbe vodom koja je dostupna biljkama pri niskoj vlažnosti zraka. Atmosfersku sušu karakterišu visoka temperatura i niska relativna vlažnost (10-20%). Ozbiljna atmosferska suša uzrokovana je kretanjem masa suhog i toplog zraka - suhim vjetrom. Zamaglica dovodi do ozbiljnih posljedica kada je suvi vjetar praćen pojavom čestica tla u zraku (oluja prašine).
Atmosferska suša, koja naglo povećava isparavanje vode s površine tla i transpiraciju, doprinosi narušavanju koordinacije brzine ulaska vode iz tla u nadzemne organe i njenom gubitku od strane biljke, kao rezultat toga, biljka vene. . Međutim, uz dobar razvoj korijenskog sistema, atmosferska suša ne nanosi mnogo štete biljkama ako temperatura ne prelazi granicu koju biljke tolerišu. Dugotrajna atmosferska suša u nedostatku kiše dovodi do suše tla, što je opasnije za biljke.
Otpornost na sušu je posledica genetski uslovljene prilagodljivosti biljaka uslovima staništa, kao i prilagođavanja na nedostatak vode. Otpornost na sušu se izražava u sposobnosti biljaka da izdrže značajnu dehidraciju zbog razvoja visokog vodnog potencijala tkiva uz funkcionalno očuvanje ćelijskih struktura, kao i zbog adaptivnih morfoloških osobina stabljike, listova, generativnih organa, koji povećavaju njihovu izdržljivost, toleranciju na efekte dugotrajne suše.
Vrste biljaka u odnosu na vodni režim
Biljke sušnih područja nazivaju se kserofiti (od grčkog xeros - suh). Sposobni su da se u procesu individualnog razvoja prilagode atmosferskoj i zemljišnoj suši. Karakteristične karakteristike kserofita su mala veličina njihove površine koja isparava, kao i mala veličina nadzemnog dijela u odnosu na podzemni. Kserofiti su obično biljke ili zakržljali grmovi. Podijeljeni su u nekoliko tipova. Predstavljamo klasifikaciju kserofita prema P. A. Genkelu.
Sukulenti su vrlo otporni na pregrijavanje i otporni na dehidraciju, tokom suše im ne nedostaje vode, jer je sadrže veliku količinu i polako je troše. Njihov korijenski sistem je razgranat u svim smjerovima u gornjim slojevima tla, zbog čega biljke u kišnim periodima brzo upijaju vodu. To su kaktusi, aloja, stonecrop, mladi.
Eukserofiti su biljke otporne na toplinu koje dobro podnose sušu. U ovu grupu spadaju stepske biljke kao što su veronika siva, dlakava astra, plavi pelin, lubenica kolocinta, devin trn i dr. Imaju nisku transpiraciju, visok osmotski pritisak, citoplazma je vrlo elastična i viskozna, korenov sistem je veoma razgranat, a njegov masa se postavlja u gornji sloj zemlje (50-60 cm). Ovi kserofiti su sposobni odbaciti lišće, pa čak i cijele grane.
Hemikserofiti ili polukserofiti su biljke koje ne podnose dehidraciju i pregrijavanje. Viskoznost i elastičnost njihovog protoplasta je neznatna, odlikuje se visokom transpiracijom, dubokim korijenskim sistemom koji može doći do podzemne vode, što osigurava nesmetano snabdijevanje vodom biljke. U ovu grupu spadaju kadulja, obični rezač itd.
Stipakserofshpy su perjanica, tyrsa i druge uskolisne stepske trave. Otporne su na pregrijavanje, dobro iskorištavaju vlagu kratkotrajnih kiša. Izdrži samo kratkotrajni nedostatak vode u tlu.
Poikilokserofiti su biljke koje ne regulišu svoj vodni režim. To su uglavnom lišajevi, koji se mogu osušiti do zračno suhog stanja i ponovo postati aktivni nakon kiše.
Higrofiti (od grčkog hihros - mokar). Biljke koje pripadaju ovoj grupi nemaju adaptacije koje ograničavaju potrošnju vode. Higrofite karakteriziraju relativno velike veličine ćelija, ljuska tankih stijenki, slabo odrveni zidovi posuda, vlakna drveta i lika, tanka kutikula i blago zadebljali vanjski zidovi epiderme, veliki puči i mali broj njih po jedinici površine, velika lisna ploča, slabo razvijena mehanička tkiva, rijetka mreža vena na listu, velika kutikularna transpiracija, duga stabljika, nerazvijen korijenski sistem. Po strukturi, higrofiti se približavaju biljkama otpornim na sjenu, ali imaju osebujnu higromorfnu strukturu. Lagani nedostatak vode u tlu uzrokuje brzo venuće higrofita. Osmotski pritisak ćelijskog soka u njima je nizak. To uključuje mannik, divlji ruzmarin, brusnice, odojke.
Prema uslovima rasta i strukturnim karakteristikama, biljke sa listovima koji su delimično ili potpuno uronjeni u vodu ili plutaju na njenoj površini, koje se nazivaju hidrofiti, veoma su bliske higrofitima.
Mezofiti (od grčkog mesos - srednji, srednji). Biljke ove ekološke grupe rastu u uslovima dovoljne vlage. Osmotski pritisak ćelijskog soka u mezofitima je 1-1,5 hiljada kPa. Lako će uvenuti. U mezofite spada većina livadskih trava i mahunarki - puzavica, lisica lisica, livadska timoteja, plava lucerka itd. Od ratarskih kultura, tvrde i meke pšenice, kukuruza, zobi, graška, soje, šećerne repe, konoplje, gotovo sve voće (sa osim badema, grožđa), mnoge povrtarske kulture (mrkva, paradajz, itd.).
Transpirirajući organi - listovi se odlikuju značajnom plastičnošću; u zavisnosti od uslova uzgoja u njihovoj strukturi uočavaju se prilično velike razlike. Čak i listovi iste biljke s različitim vodosnabdijevanjem i osvjetljenjem imaju razlike u strukturi. Utvrđeni su određeni uzorci u strukturi listova, ovisno o njihovoj lokaciji na biljci.
V. R. Zalensky je otkrio promjene u anatomskoj strukturi listova po slojevima. Utvrdio je da listovi gornjeg sloja pokazuju redovite promjene u smjeru povećanja kseromorfizma, odnosno formiraju se strukture koje povećavaju otpornost ovih listova na sušu. Listovi koji se nalaze u gornjem dijelu stabljike uvijek se razlikuju od donjih, i to: što je list viši na stabljici, to je manja veličina njegovih ćelija, to je veći broj stomata i manja njihova veličina. što je veći broj dlaka po jedinici površine, što je mreža vaskularnih snopova gušća, to je jače palisadno tkivo razvijeno. Svi ovi znakovi karakteriziraju kserofiliju, odnosno stvaranje struktura koje doprinose povećanju otpornosti na sušu.
Fiziološke karakteristike su povezane i sa određenom anatomskom strukturom, odnosno: gornji listovi se odlikuju većom sposobnošću asimilacije i intenzivnijom transpiracijom. Koncentracija soka u gornjim listovima je također veća, pa zbog toga gornji listovi mogu odvlačiti vodu iz donjih listova, što dovodi do sušenja i odumiranja donjih listova. Struktura organa i tkiva koja povećava otpornost biljaka na sušu naziva se kseromorfizam. Prepoznatljive karakteristike u strukturi listova gornjeg sloja objašnjavaju se činjenicom da se razvijaju u uvjetima donekle otežane vodoopskrbe.
Formiran je složen sistem anatomskih i fizioloških adaptacija za izjednačavanje ravnoteže između dotoka i odliva vode u biljci. Takve adaptacije se primjećuju kod kserofita, higrofita, mezofita.
Rezultati istraživanja su pokazali da adaptivna svojstva biljnih oblika otpornih na sušu nastaju pod uticajem uslova njihovog postojanja.
ZAKLJUČAK
Zadivljujući sklad žive prirode, njeno savršenstvo stvara sama priroda: borba za opstanak. Oblici adaptacije kod biljaka i životinja su beskrajno raznoliki. Čitav životinjski i biljni svijet se od svog nastanka usavršavao putem svrsishodnog prilagođavanja životnim uvjetima: vodi, zraku, sunčevoj svjetlosti, gravitaciji itd.
LITERATURA
1. Volodko I.K. "Mikroelementi i otpornost biljaka na nepovoljne uslove", Minsk, Nauka i tehnologija, 1983.
2. Goryshina T.K. "Ekologija biljaka"", uč. Priručnik za univerzitete, Moskva, V. škola, 1979.
3. Prokofjev A.A. "Problemi otpornosti biljaka na sušu", Moskva, Nauka, 1978.
4. Sergejeva K.A. "" Fiziološke i biohemijske osnove zimske otpornosti drvenastih biljaka ", Moskva, Nauka, 1971.
5. Kultiasov I.M. Ekologija biljaka. - M.: Izdavačka kuća Moskovskog univerziteta, 1982
Prilagodljivost ontogeneze biljaka na uslove sredine rezultat je njihovog evolucionog razvoja (varijabilnost, nasljednost, selekcija). Tokom filogeneze svake biljne vrste, u procesu evolucije, razvile su se određene potrebe pojedinca za uslovima postojanja i prilagodljivosti ekološkoj niši koju zauzima. Tolerantnost na vlagu i hlad, otpornost na toplotu, hladnoću i druge ekološke karakteristike pojedinih biljnih vrsta formirane su tokom evolucije kao rezultat dugotrajnog izlaganja odgovarajućim uslovima. Dakle, biljke koje vole toplinu i biljke kratkog dana karakteristične su za južne geografske širine, manje zahtjevne za toplinu i biljke dugog dana - za sjeverne.
U prirodi, u jednoj geografskoj regiji, svaka biljna vrsta zauzima ekološku nišu koja odgovara njenim biološkim karakteristikama: voli vlagu - bliže vodenim tijelima, tolerantna na sjenu - pod krošnjom šume, itd. Nasljednost biljaka se formira pod utjecajem određenih uslova životne sredine. Važni su i vanjski uslovi ontogeneze biljaka.
U većini slučajeva, biljke i usjevi (zasadi) poljoprivrednih kultura, doživljavajući djelovanje određenih nepovoljnih faktora, pokazuju otpor prema njima kao rezultat prilagođavanja uvjetima postojanja koji su se povijesno razvijali, što je primijetio K. A. Timiryazev.
1. Osnovne životne sredine.
Prilikom proučavanja životne sredine (staništa biljaka i životinja i ljudskih proizvodnih aktivnosti) izdvajaju se sledeće glavne komponente: vazdušna sredina; vodena sredina (hidrosfera); fauna (ljudi, domaće i divlje životinje, uključujući ribe i ptice); flora (kultivisane i samonikle biljke, uključujući i one koje rastu u vodi), tlo (vegetacijski sloj), podzemlje (gornji dio zemljine kore, unutar kojeg je moguće rudarenje); klimatsko i akustičko okruženje.
Vazdušna sredina može biti eksterna, u kojoj većina ljudi provodi manji dio svog vremena (do 10-15%), interna proizvodna (u njoj osoba provodi do 25-30% vremena) i unutrašnja stambena, gdje ljudi ostaju većinu vremena (do 60 -70% ili više).
Vanjski vazduh na površini zemlje sadrži po zapremini: 78,08% azota; 20,95% kiseonika; 0,94% inertnih plinova i 0,03% ugljičnog dioksida. Na visini od 5 km sadržaj kiseonika ostaje isti, dok se azota povećava na 78,89%. Često zrak u blizini površine zemlje ima razne nečistoće, posebno u gradovima: tamo sadrži više od 40 sastojaka koji su stranci prirodnom zračnom okruženju. Unutrašnji vazduh u stanovima, po pravilu, ima
povećan sadržaj ugljičnog dioksida, a unutrašnji zrak industrijskih prostorija obično sadrži nečistoće, čija je priroda određena tehnologijom proizvodnje. Među plinovima se oslobađa vodena para, koja ulazi u atmosferu kao rezultat isparavanja sa Zemlje. Najveći dio (90%) koncentrisan je u najnižem pet-kilometarskom sloju atmosfere, s visinom se njegova količina vrlo brzo smanjuje. Atmosfera sadrži mnogo prašine koja tamo stiže sa površine Zemlje i dijelom iz svemira. Za vrijeme jakih valova vjetrovi podižu vodeni mlaz iz mora i okeana. Tako čestice soli iz vode dospiju u atmosferu. Kao rezultat vulkanskih erupcija, šumskih požara, industrijskih objekata itd. vazduh je zagađen produktima nepotpunog sagorevanja. Najviše prašine i drugih nečistoća nalazi se u prizemnom sloju vazduha. Čak i nakon kiše, 1 cm sadrži oko 30 hiljada čestica prašine, a po suhom ih je nekoliko puta više po suhom vremenu.
Sve ove sitne nečistoće utiču na boju neba. Molekuli gasova raspršuju kratkotalasni dio spektra sunčevog zraka, tj. ljubičastih i plavih zraka. Dakle, tokom dana je nebo plavo. A čestice nečistoća, koje su mnogo veće od molekula plina, raspršuju svjetlosne zrake gotovo svih valnih dužina. Stoga, kada je zrak prašnjav ili sadrži kapljice vode, nebo postaje bjelkasto. Na velikim visinama nebo je tamnoljubičasto, pa čak i crno.
Kao rezultat fotosinteze koja se odvija na Zemlji, vegetacija godišnje formira 100 milijardi tona organskih supstanci (oko polovine otpada na mora i okeane), dok asimilira oko 200 milijardi tona ugljičnog dioksida i ispušta oko 145 milijardi tona u okruženje. slobodnog kiseonika, veruje se da fotosintezom nastaje sav kiseonik u atmosferi. O ulozi zelenih površina u ovom ciklusu govore sledeći podaci: 1 hektar zelenih površina čisti vazduh od 8 kg ugljen-dioksida u proseku za 1 sat (200 ljudi emituje za to vreme prilikom disanja). Odraslo drvo oslobađa 180 litara kiseonika dnevno, a za pet meseci (od maja do septembra) apsorbuje oko 44 kg ugljen-dioksida.
Količina oslobođenog kisika i apsorbiranog ugljičnog dioksida ovisi o starosti zelenih površina, sastavu vrsta, gustoći sadnje i drugim faktorima.
Jednako su važne i morske biljke – fitoplankton (uglavnom alge i bakterije), koje fotosintezom oslobađaju kisik.
Vodeni okoliš uključuje površinske i podzemne vode. Površinske vode su uglavnom koncentrisane u okeanu, sa sadržajem od 1 milijardu 375 miliona kubnih kilometara - oko 98% sve vode na Zemlji. Površina okeana (vodno područje) je 361 milion kvadratnih kilometara. To je oko 2,4 puta više od površine zemlje - teritorija koja zauzima 149 miliona kvadratnih kilometara. Voda u okeanu je slana, a najveći dio (više od 1 milijarde kubnih kilometara) zadržava konstantan salinitet od oko 3,5% i temperaturu od oko 3,7°C. Primjetne razlike u salinitetu i temperaturi uočavaju se gotovo isključivo na površini sloju vode, kao iu rubnim i posebno u Sredozemnom moru. Sadržaj otopljenog kisika u vodi značajno opada na dubini od 50-60 metara.
Podzemne vode mogu biti slane, bočate (nižeg saliniteta) i svježe; postojeće geotermalne vode imaju povišenu temperaturu (više od 30ºC).
Za proizvodne aktivnosti čovječanstva i potrebe njegovih domaćinstava potrebna je slatka voda, čija količina iznosi samo 2,7% ukupne količine vode na Zemlji, a njen vrlo mali udio (samo 0,36%) je dostupan na mjestima koja su lako dostupni za vađenje. Većina slatke vode nalazi se u snijegu i slatkovodnim santima leda koji se nalaze u područjima prvenstveno u Antarktičkom krugu.
Godišnji svjetski riječni otjecanje slatke vode iznosi 37,3 hiljade kubnih kilometara. Osim toga, može se koristiti i dio podzemne vode od 13 hiljada kubnih kilometara. Nažalost, najveći dio riječnog toka u Rusiji, koji iznosi oko 5.000 kubnih kilometara, pada na rubne i slabo naseljene sjeverne teritorije.
Klimatsko okruženje je važan faktor koji determiniše razvoj različitih vrsta flore i faune i njenu plodnost. Karakteristična karakteristika Rusije je da većina njene teritorije ima mnogo hladniju klimu nego u drugim zemljama.
Sve razmatrane komponente životne sredine su uključene u
BIOSFERA: Zemljina školjka, uključujući dio atmosfere, hidrosferu i gornji dio litosfere, koji su međusobno povezani složenim biohemijskim ciklusima migracije materije i energije, geološka ljuska Zemlje, naseljena živim organizmima. Gornja granica života biosfere ograničena je intenzivnom koncentracijom ultraljubičastih zraka; niža - visoka temperatura unutrašnjosti zemlje (preko 100`C). Njegove ekstremne granice dostižu samo niži organizmi - bakterije.
Adaptacija (prilagođavanje) biljke specifičnim uslovima sredine obezbeđuje se fiziološkim mehanizmima (fiziološka adaptacija), au populaciji organizama (vrste) - mehanizmima genetske varijabilnosti, nasleđa i selekcije (genetska adaptacija). Faktori okoline se mogu mijenjati redovno i nasumično. Redovno promjenjivi uvjeti okoline (promjena godišnjih doba) razvijaju kod biljaka genetsku adaptaciju na ove uslove.
U prirodnim uslovima rasta ili uzgoja neke vrste, u toku svog rasta i razvoja, često doživljavaju uticaj nepovoljnih faktora sredine, među kojima su temperaturne fluktuacije, suša, prekomerna vlaga, zaslanjenost zemljišta i dr. sposobnost prilagođavanja promenljivim uslovima.uslovi sredine u granicama koje određuje njen genotip. Što je veća sposobnost biljke da mijenja metabolizam u skladu sa okruženjem, to je veća brzina reakcije ove biljke i bolja je sposobnost prilagođavanja. Ovo svojstvo razlikuje otporne sorte poljoprivrednih kultura. Blage i kratkotrajne promjene faktora sredine u pravilu ne dovode do značajnijih poremećaja fizioloških funkcija biljaka, što je posljedica njihove sposobnosti da održavaju relativno stabilno stanje u promjenjivim uvjetima okoline, odnosno održavaju homeostazu. Međutim, oštri i dugotrajni udari dovode do poremećaja mnogih funkcija biljke, a često i do njene smrti.
Pod uticajem nepovoljnih uslova, smanjenje fizioloških procesa i funkcija može dostići kritične nivoe koji ne obezbeđuju realizaciju genetskog programa ontogeneze, narušava se energetski metabolizam, regulacioni sistemi, metabolizam proteina i druge vitalne funkcije biljnog organizma. Kada je biljka izložena nepovoljnim faktorima (stresorima), u njoj nastaje stresno stanje, odstupanje od norme - stres. Stres je opća nespecifična adaptivna reakcija organizma na djelovanje bilo kojeg štetnog faktora. Postoje tri glavne grupe faktora koji izazivaju stres kod biljaka: fizički - nedovoljna ili prekomjerna vlažnost, svjetlost, temperatura, radioaktivno zračenje, mehanički stres; hemikalije - soli, gasovi, ksenobiotici (herbicidi, insekticidi, fungicidi, industrijski otpad, itd.); biološki - oštećenja od patogena ili štetočina, konkurencija s drugim biljkama, utjecaj životinja, cvjetanje, sazrijevanje plodova.
Reakcije na nepovoljne faktore okoline samo su pod određenim uslovima štetne za žive organizme, au većini slučajeva imaju adaptivnu vrijednost. Stoga je ove odgovore Selye nazvao "sindromom opšte adaptacije". U kasnijim radovima koristio je pojmove "stres" i "sindrom generalne adaptacije" kao sinonime.
Adaptacija- ovo je genetski determinisani proces formiranja zaštitnih sistema koji obezbeđuju povećanje stabilnosti i tok ontogeneze u za nju nepovoljnim uslovima.
Adaptacija je jedan od najvažnijih mehanizama koji povećava stabilnost biološkog sistema, pa i biljnog organizma, u promenjenim uslovima postojanja. Što je organizam bolje prilagođen nekom faktoru, to je otporniji na njegove fluktuacije.
Genotipski određena sposobnost organizma da mijenja metabolizam u određenim granicama, u zavisnosti od djelovanja vanjske sredine, naziva se brzina reakcije. Kontroliše ga genotip i karakterističan je za sve žive organizme. Većina modifikacija koje se dešavaju u granicama reakcione norme su od adaptivnog značaja. Odgovaraju promjenama u staništu i omogućavaju bolji opstanak biljaka u promjenjivim uvjetima okoline. U tom smislu, takve modifikacije su od evolucijske važnosti. Termin "brzina reakcije" uveo je V.L. Johansen (1909).
Što je veća sposobnost vrste ili varijeteta da se modificira u skladu sa okolinom, to je šira brzina njene reakcije i veća je sposobnost prilagođavanja. Ovo svojstvo razlikuje otporne sorte poljoprivrednih kultura. U pravilu, blage i kratkotrajne promjene faktora okoliša ne dovode do značajnijih narušavanja fizioloških funkcija biljaka. To je zbog njihove sposobnosti da održe relativnu dinamičku ravnotežu unutrašnjeg okruženja i stabilnost osnovnih fizioloških funkcija u promjenjivom vanjskom okruženju. Istovremeno, oštri i dugotrajni udari dovode do poremećaja mnogih funkcija biljke, a često i do njene smrti.
Adaptacija uključuje sve procese i adaptacije (anatomske, morfološke, fiziološke, bihevioralne, itd.) koji povećavaju stabilnost i doprinose opstanku vrste.
1.Anatomske i morfološke adaptacije. Kod nekih predstavnika kserofita, dužina korijenskog sistema doseže nekoliko desetina metara, što biljci omogućava da koristi podzemne vode i ne doživi nedostatak vlage u uvjetima tla i atmosferske suše. Kod ostalih kserofita, prisustvo debele kutikule, pubescencija listova i transformacija listova u bodlje smanjuju gubitak vode, što je veoma važno u uslovima nedostatka vlage.
Zapaljene dlake i bodlje štite biljke od toga da ih životinje pojedu.
Drveće u tundri ili na visokim planinskim visinama izgleda kao zdepasto puzavo grmlje, zimi je prekriveno snijegom, koji ih štiti od jakih mrazeva.
U planinskim područjima sa velikim dnevnim temperaturnim kolebanjima, biljke često imaju oblik spljoštenih jastuka sa gusto raspoređenim brojnim stabljikama. To vam omogućava da zadržite vlagu unutar jastuka i relativno ujednačenu temperaturu tokom cijelog dana.
U močvarnim i vodenim biljkama formira se poseban vazdušni parenhim (aerenhim) koji je rezervoar vazduha i olakšava disanje biljnih delova potopljenih u vodu.
2. Fiziološke i biohemijske adaptacije. U sukulentima, adaptacija za uzgoj u pustinjskim i polupustinjskim uslovima je asimilacija CO 2 tokom fotosinteze duž CAM puta. Ove biljke imaju zatvorene puške tokom dana. Tako postrojenje čuva unutrašnje rezerve vode od isparavanja. U pustinjama, voda je glavni faktor koji ograničava rast biljaka. Stoma se otvara noću i u to vrijeme CO 2 ulazi u fotosintetska tkiva. Naknadno uključivanje CO2 u fotosintetski ciklus događa se danju već sa zatvorenim stomama.
Fiziološke i biohemijske adaptacije uključuju mogućnost otvaranja i zatvaranja stomata, u zavisnosti od spoljašnjih uslova. Sinteza u stanicama apscizinske kiseline, prolina, zaštitnih proteina, fitoaleksina, fitoncida, povećanje aktivnosti enzima koji suzbijaju oksidativni razgradnju organskih tvari, nakupljanje šećera u stanicama i niz drugih promjena u metabolizmu doprinose povećanje otpornosti biljaka na nepovoljne uslove okoline.
Istu biokemijsku reakciju može provesti nekoliko molekularnih oblika istog enzima (izoenzima), dok svaka izoforma ispoljava katalitičku aktivnost u relativno uskom rasponu nekih parametara okoline, kao što je temperatura. Prisustvo brojnih izoenzima omogućava biljci da izvede reakciju u mnogo širem rasponu temperatura, u poređenju sa svakim pojedinačnim izoenzima. To omogućava biljci da uspješno obavlja vitalne funkcije u promjenjivim temperaturnim uvjetima.
3. Adaptacije ponašanja, odnosno izbjegavanje štetnog faktora. Primjer su efemera i efemeroidi (mak, zvijezda, krokusi, tulipani, snježne kapljice). Oni prolaze kroz cijeli ciklus svog razvoja u proljeće 1,5-2 mjeseca, čak i prije početka vrućine i suše. Tako na neki način odlaze, odnosno izbjegavaju da padnu pod utjecaj stresora. Na sličan način, ranozrele sorte poljoprivrednih kultura formiraju usjev prije pojave nepovoljnih sezonskih pojava: kolovozne magle, kiše, mrazeva. Stoga je odabir mnogih poljoprivrednih kultura usmjeren na stvaranje ranih zrelih sorti. Višegodišnje biljke prezimljuju kao rizomi i lukovice u tlu pod snijegom, što ih štiti od smrzavanja.
Adaptacija biljaka na nepovoljne faktore odvija se istovremeno na više nivoa regulacije - od jedne ćelije do fitocenoze. Što je viši nivo organizacije (ćelija, organizam, populacija), veći je broj mehanizama koji su istovremeno uključeni u adaptaciju biljaka na stres.
Regulacija metaboličkih i adaptivnih procesa unutar ćelije vrši se uz pomoć sistema: metaboličkog (enzimskog); genetski; membrana. Ovi sistemi su usko povezani. Dakle, svojstva membrana zavise od aktivnosti gena, a diferencijalna aktivnost samih gena je pod kontrolom membrana. Sinteza enzima i njihova aktivnost kontrolišu se na genetskom nivou, a istovremeno enzimi regulišu metabolizam nukleinskih kiselina u ćeliji.
Na nivo organizmaćelijskim mehanizmima adaptacije dodaju se novi, koji odražavaju interakciju organa. U nepovoljnim uvjetima biljke stvaraju i zadržavaju toliki broj plodnih elemenata koji su u dovoljnim količinama opskrbljeni potrebnim tvarima za formiranje punopravnog sjemena. Na primjer, u cvatovima kultivisanih žitarica i u krošnjama voćaka, u nepovoljnim uvjetima može otpasti više od polovine položenih jajnika. Takve promjene su zasnovane na kompetitivnim odnosima između organa za fiziološki aktivne i hranljive materije.
U uslovima stresa, procesi starenja i opadanja donjih listova se naglo ubrzavaju. U isto vrijeme, tvari potrebne za biljke prelaze iz njih u mlade organe, odgovarajući na strategiju preživljavanja organizma. Zahvaljujući recikliranju nutrijenata iz donjih listova, mlađi, gornji listovi, ostaju održivi.
Postoje mehanizmi regeneracije izgubljenih organa. Na primjer, površina rane je prekrivena sekundarnim integumentarnim tkivom (periderm rane), rana na deblu ili grani se zacjeljuje uljevima (kalusima). Gubitkom vršnog izdanka, u biljkama se bude uspavani pupoljci i intenzivno se razvijaju bočni izdanci. Proljetna obnova lišća umjesto opalog u jesen također je primjer prirodne regeneracije organa. Regeneracija kao biološki uređaj koji obezbeđuje vegetativno razmnožavanje biljaka segmentima korena, rizoma, steljke, stabljike i lisnih reznica, izolovanih ćelija, pojedinačnih protoplasta, od velikog je praktičnog značaja za biljnu proizvodnju, voćarstvo, šumarstvo, ukrasno baštovanstvo itd.
Hormonski sistem je takođe uključen u procese zaštite i adaptacije na biljnom nivou. Na primjer, pod utjecajem nepovoljnih uvjeta u biljci, naglo se povećava sadržaj inhibitora rasta: etilena i apscisne kiseline. Smanjuju metabolizam, inhibiraju procese rasta, ubrzavaju starenje, opadanje organa i prelazak biljke u stanje mirovanja. Inhibicija funkcionalne aktivnosti pod stresom pod uticajem inhibitora rasta je karakteristična reakcija za biljke. Istovremeno se smanjuje sadržaj stimulansa rasta u tkivima: citokinina, auksina i giberelina.
Na nivo populacije dodaje se selekcija, što dovodi do pojave prilagođenijih organizama. Mogućnost selekcije određena je postojanjem intrapopulacijske varijabilnosti otpornosti biljaka na različite faktore sredine. Primjer intrapopulacijske varijabilnosti otpornosti može biti neprijatan izgled sadnica na zaslanjenom tlu i povećanje varijacije u vremenu klijanja s povećanjem djelovanja stresora.
Vrsta u savremenom pogledu sastoji se od velikog broja biotipova - manjih ekoloških jedinica, genetski identičnih, ali pokazuju različitu otpornost na faktore sredine. U različitim uslovima nisu svi biotipovi podjednako vitalni, a kao rezultat konkurencije ostaju samo oni od njih koji najbolje ispunjavaju date uslove. Odnosno, otpor populacije (varijeteta) na određeni faktor je određen otpornošću organizama koji čine populaciju. Otporne sorte u svom sastavu imaju skup biotipova koji pružaju dobru produktivnost čak iu nepovoljnim uvjetima.
Istovremeno, u procesu dugotrajnog uzgoja, sastav i omjer biotipova u populaciji se mijenja u sortama, što utiče na produktivnost i kvalitet sorte, često ne na bolje.
Dakle, adaptacija uključuje sve procese i adaptacije koje povećavaju otpornost biljaka na nepovoljne uslove okoline (anatomske, morfološke, fiziološke, biohemijske, bihevioralne, populacione itd.)
Ali da biste odabrali najefikasniji način adaptacije, glavna stvar je vrijeme tokom kojeg se tijelo mora prilagoditi novim uvjetima.
Uz iznenadno djelovanje ekstremnog faktora, reakcija se ne može odgoditi, ona mora uslijediti odmah kako bi se isključila nepovratna oštećenja biljke. Kod dugoročnih uticaja male sile, adaptivna prestrojavanja se javljaju postepeno, dok se izbor mogućih strategija povećava.
U tom smislu, postoje tri glavne strategije prilagođavanja: evolucijski, ontogenetski i hitno. Zadatak strategije je efikasno korištenje raspoloživih resursa za postizanje glavnog cilja – opstanak organizma pod stresom. Strategija adaptacije ima za cilj održavanje strukturnog integriteta vitalnih makromolekula i funkcionalne aktivnosti ćelijskih struktura, održavanje sistema regulacije vitalne aktivnosti i snabdevanje biljaka energijom.
Evolucijske ili filogenetske adaptacije(filogenija - razvoj biološke vrste u vremenu) - to su adaptacije koje nastaju tokom evolucijskog procesa na osnovu genetskih mutacija, selekcije i nasljeđuju se. Oni su najpouzdaniji za preživljavanje biljaka.
Svaka vrsta biljaka u procesu evolucije razvila je određene potrebe za uslovima postojanja i prilagodljivosti ekološkoj niši koju zauzima, stabilnom adaptacijom organizma na okolinu. Tolerancija na vlagu i hladovinu, otpornost na toplotu, otpornost na hladnoću i druge ekološke karakteristike pojedinih biljnih vrsta formirane su kao rezultat dugotrajnog delovanja relevantnih uslova. Tako su biljke koje vole toplinu i biljke kratkog dana karakteristične za južne geografske širine, a manje zahtjevne za toplinom i biljke dugog dana karakteristične su za sjeverne geografske širine. Poznate su brojne evolucijske adaptacije biljaka kserofita na sušu: ekonomično korištenje vode, dubok korijenski sistem, opadanje lišća i prelazak u stanje mirovanja i druge adaptacije.
S tim u vezi sorte poljoprivrednih biljaka pokazuju otpornost upravo na one faktore okoline protiv kojih se vrši uzgoj i selekcija proizvodnih oblika. Ako se selekcija odvija u nizu uzastopnih generacija u pozadini stalnog utjecaja nekog nepovoljnog faktora, tada se otpornost sorte na njega može značajno povećati. Prirodno je da su sorte koje uzgaja Istraživački institut za poljoprivredu Jugoistoka (Saratov) otpornije na sušu od sorti stvorenih u uzgojnim centrima moskovske regije. Na isti način, u ekološkim zonama sa nepovoljnim zemljišno-klimatskim uslovima formirane su otporne lokalne biljne sorte, a endemske biljne vrste otporne su na stresor koji je izražen u njihovom staništu.
Karakterizacija otpornosti sorti jare pšenice iz kolekcije Sveruskog instituta za biljnu industriju (Semenov et al., 2005.)
| Raznolikost | Porijeklo | Održivost |
| Enita | Moskva region | Srednje otporan na sušu |
| Saratovskaja 29 | Saratov region | otporan na sušu |
| Comet | Sverdlovsk region. | otporan na sušu |
| Karazino | Brazil | otporan na kiseline |
| Preludij | Brazil | otporan na kiseline |
| Kolonias | Brazil | otporan na kiseline |
| Thrintani | Brazil | otporan na kiseline |
| PPG-56 | Kazahstan | otporan na sol |
| Osh | Kirgistan | otporan na sol |
| Surkhak 5688 | Tadžikistan | otporan na sol |
| Messel | Norveška | Otporan na sol |
U prirodnom okruženju uslovi sredine se obično veoma brzo menjaju, a vreme tokom kojeg faktor stresa dostigne štetni nivo nije dovoljno za formiranje evolucionih adaptacija. U tim slučajevima biljke koriste ne trajne, već stresorom izazvane odbrambene mehanizme, čije je formiranje genetski predodređeno (definirano).
Ontogenetske (fenotipske) adaptacije nisu povezani s genetskim mutacijama i nisu naslijeđeni. Formiranje takvih adaptacija zahtijeva relativno dugo vrijeme, pa se nazivaju dugotrajnim adaptacijama. Jedan od takvih mehanizama je sposobnost brojnih biljaka da formiraju put fotosinteze tipa CAM koji štedi vodu u uslovima nedostatka vode uzrokovanog sušom, salinitetom, niskim temperaturama i drugim stresorima.
Ova adaptacija je povezana sa indukcijom ekspresije gena fosfoenolpiruvat karboksilaze, koji je u normalnim uslovima neaktivan, i gena drugih enzima CAM puta unosa CO2, sa biosintezom osmolita (prolina), uz aktivaciju antioksidansa. sistema, te sa promjenama u dnevnim ritmovima stomatalnih pokreta. Sve to dovodi do vrlo ekonomične potrošnje vode.
U ratarskim kulturama, na primjer, u kukuruzu, aerenhim je odsutan u normalnim uslovima uzgoja. Ali u uslovima poplave i nedostatka kiseonika u tkivima u korenu, neke od ćelija primarnog korteksa korena i stabljike umiru (apoptoza, ili programirana ćelijska smrt). Na njihovom mjestu formiraju se šupljine kroz koje se kisik prenosi iz nadzemnog dijela biljke do korijenskog sistema. Signal za smrt ćelije je sinteza etilena.
Hitna adaptacija javlja se brzim i intenzivnim promjenama životnih uslova. Zasnovan je na formiranju i funkcionisanju sistema zaštite od udara. Sistemi odbrane od šoka uključuju, na primjer, sistem proteina toplotnog šoka, koji se formira kao odgovor na brzo povećanje temperature. Ovi mehanizmi obezbeđuju kratkoročne uslove preživljavanja pod dejstvom štetnog faktora i na taj način stvaraju preduslove za formiranje pouzdanijih dugoročnih specijalizovanih mehanizama adaptacije. Primjer specijalizovanih mehanizama adaptacije je novo formiranje antifriz proteina na niskim temperaturama ili sinteza šećera tokom prezimljavanja ozimih usjeva. U isto vrijeme, ako štetni učinak faktora premašuje zaštitne i reparativne sposobnosti tijela, tada neizbježno dolazi do smrti. U tom slučaju organizam umire u fazi hitne ili u fazi specijalizirane adaptacije, ovisno o intenzitetu i trajanju djelovanja ekstremnog faktora.
Razlikovati specifično i nespecifičan (opći) reakcije biljaka na stresore.
Nespecifične reakcije ne zavise od prirode faktora koji djeluje. Isti su pod dejstvom visokih i niskih temperatura, manjka ili viška vlage, visoke koncentracije soli u tlu ili štetnih gasova u vazduhu. U svim slučajevima povećava se propusnost membrana u biljnim stanicama, poremećeno je disanje, povećava se hidrolitička razgradnja tvari, povećava se sinteza etilena i apscizinske kiseline, inhibira se dioba i produljenje stanica.
U tabeli je prikazan kompleks nespecifičnih promjena koje nastaju u biljkama pod utjecajem različitih faktora okoline.
Promene fizioloških parametara u biljkama pod uticajem stresnih uslova (prema G.V., Udovenko, 1995.)
| Opcije | Priroda promjene parametara pod uvjetima | |||
| suše | salinitet | visoke temperature | niske temperature | |
| Koncentracija jona u tkivima | raste | raste | raste | raste |
| Aktivnost vode u ćeliji | Padati | Padati | Padati | Padati |
| Osmotski potencijal ćelije | raste | raste | raste | raste |
| Kapacitet zadržavanja vode | raste | raste | raste | — |
| Nestašica vode | raste | raste | raste | — |
| Permeabilnost protoplazme | raste | raste | raste | — |
| Stopa transpiracije | Padati | Padati | raste | Padati |
| Efikasnost transpiracije | Padati | Padati | Padati | Padati |
| Energetska efikasnost disanja | Padati | Padati | Padati | — |
| Intenzitet disanja | raste | raste | raste | — |
| Fotofosforilacija | Smanjuje | Smanjuje | — | Smanjuje |
| Stabilizacija nuklearne DNK | raste | raste | raste | raste |
| Funkcionalna aktivnost DNK | Smanjuje | Smanjuje | Smanjuje | Smanjuje |
| Koncentracija prolina | raste | raste | raste | — |
| Sadržaj proteina rastvorljivih u vodi | raste | raste | raste | raste |
| Sintetičke reakcije | Potisnuto | Potisnuto | Potisnuto | Potisnuto |
| Upijanje jona od strane korijena | Potisnuto | Potisnuto | Potisnuto | Potisnuto |
| Transport materija | Depresivan | Depresivan | Depresivan | Depresivan |
| Koncentracija pigmenta | Padati | Padati | Padati | Padati |
| ćelijska dioba | usporava | usporava | — | — |
| Cell stretch | Potisnuto | Potisnuto | — | — |
| Broj voćnih elemenata | Smanjeno | Smanjeno | Smanjeno | Smanjeno |
| Starenje organa | Ubrzano | Ubrzano | Ubrzano | — |
| biološka žetva | Degradiran | Degradiran | Degradiran | Degradiran |
Na osnovu podataka u tabeli može se vidjeti da je otpornost biljaka na više faktora praćena jednosmjernim fiziološkim promjenama. To daje razlog za vjerovanje da povećanje otpornosti biljaka na jedan faktor može biti praćeno povećanjem otpornosti na drugi. Ovo je potvrđeno eksperimentima.
Eksperimenti na Institutu za fiziologiju bilja Ruske akademije nauka (Vl. V. Kuznjecov i drugi) pokazali su da kratkotrajna termička obrada biljaka pamuka prati povećanje njihove otpornosti na kasniju salinizaciju. A prilagođavanje biljaka na slanost dovodi do povećanja njihove otpornosti na visoke temperature. Toplotni šok povećava sposobnost biljaka da se prilagode narednoj suši i, obrnuto, u procesu suše raste otpornost organizma na visoke temperature. Kratkotrajno izlaganje visokim temperaturama povećava otpornost na teške metale i UV-B zračenje. Prethodna suša pogoduje opstanku biljaka u uslovima saliniteta ili hladnoće.
Proces povećanja otpornosti organizma na dati faktor okoline kao rezultat adaptacije na faktor drugačije prirode naziva se unakrsna adaptacija.
Za proučavanje općih (nespecifičnih) mehanizama rezistencije od velikog je interesa odgovor biljaka na faktore koji uzrokuju nedostatak vode u biljkama: salinitet, sušu, niske i visoke temperature i neke druge. Na nivou cijelog organizma, sve biljke na isti način reaguju na nedostatak vode. Karakterizira ga inhibicija rasta izdanaka, pojačan rast korijenskog sistema, sinteza apscizinske kiseline i smanjenje provodljivosti stomata. Nakon nekog vremena, donji listovi brzo stare i opaža se njihovo odumiranje. Sve ove reakcije imaju za cilj smanjenje potrošnje vode smanjenjem površine isparavanja, kao i povećanjem upijajuće aktivnosti korijena.
Specifične reakcije su reakcije na djelovanje bilo kojeg faktora stresa. Dakle, fitoaleksini (supstance sa antibiotskim svojstvima) se sintetiziraju u biljkama kao odgovor na kontakt sa patogenima (patogeni).
Specifičnost ili nespecifičnost odgovora podrazumijeva, s jedne strane, odnos biljke prema različitim stresorima, a s druge strane karakteristične reakcije biljaka različitih vrsta i sorti na isti stresor.
Ispoljavanje specifičnih i nespecifičnih odgovora biljaka zavisi od jačine stresa i brzine njegovog razvoja. Specifične reakcije se češće javljaju ako se stres sporo razvija, a tijelo ima vremena da se obnovi i prilagodi mu. Nespecifične reakcije obično se javljaju uz kraće i jače djelovanje stresora. Funkcioniranje nespecifičnih (općih) mehanizama otpora omogućava biljci da izbjegne velike energetske troškove za formiranje specijaliziranih (specifičnih) mehanizama adaptacije kao odgovor na bilo koje odstupanje od norme u njihovim životnim uvjetima.
Otpornost biljaka na stres zavisi od faze ontogeneze. Najstabilnije biljke i biljni organi u stanju mirovanja: u obliku sjemena, lukovica; drvenaste trajnice - u stanju dubokog mirovanja nakon opadanja listova. Biljke su najosjetljivije u mladosti, jer se u stresnim uslovima prvenstveno oštećuju procesi rasta. Drugi kritični period je period formiranja gameta i oplodnje. Dejstvo stresa u ovom periodu dovodi do smanjenja reproduktivne funkcije biljaka i smanjenja prinosa.
Ako se stresni uslovi ponavljaju i imaju mali intenzitet, onda doprinose otvrdnjavanju biljaka. Ovo je osnova za metode za povećanje otpornosti na niske temperature, toplotu, salinitet i povećan sadržaj štetnih gasova u vazduhu.
Pouzdanost biljnog organizma određena je njegovom sposobnošću da spriječi ili eliminiše kvarove na različitim nivoima biološke organizacije: molekularnoj, subćelijskoj, ćelijskoj, tkivnoj, organskoj, organskoj i populacijskoj.
Kako bi se spriječili poremećaji u životu biljaka pod utjecajem nepovoljnih faktora, principi redundantnost, heterogenost funkcionalno ekvivalentnih komponenti, sistemi za popravku izgubljenih konstrukcija.
Redundantnost struktura i funkcionalnosti jedan je od glavnih načina da se osigura pouzdanost sistema. Redundancija i redundantnost imaju višestruke manifestacije. Na subćelijskom nivou, rezervisanje i umnožavanje genetskog materijala doprinosi povećanju pouzdanosti biljnog organizma. To se postiže, na primjer, dvostrukom spiralom DNK, povećanjem ploidnosti. Pouzdanost funkcionisanja biljnog organizma u promjenjivim uvjetima održava se i zbog prisustva različitih molekula RNK glasnika i formiranja heterogenih polipeptida. To uključuje izoenzime koji kataliziraju istu reakciju, ali se razlikuju po svojim fizičko-hemijskim svojstvima i stabilnosti molekularne strukture u promjenjivim uvjetima okoline.
Na ćelijskom nivou, primjer redundancije je višak ćelijskih organela. Tako je utvrđeno da je dio raspoloživih hloroplasta dovoljan da biljci obezbijedi produkte fotosinteze. Preostali hloroplasti, takoreći, ostaju u rezervi. Isto važi i za ukupan sadržaj hlorofila. Redundantnost se također manifestira u velikoj akumulaciji prekursora za biosintezu mnogih jedinjenja.
Na nivou organizma, princip redundancije se izražava u formiranju i polaganju u različito vrijeme više izdanaka, cvjetova, klasića nego što je potrebno za smjenu generacija, u ogromnoj količini polena, sjemenki, sjemenki.
Na nivou populacije, princip redundancije se manifestuje u velikom broju jedinki koje se razlikuju po otpornosti na određeni faktor stresa.
Sistemi za popravke također rade na različitim nivoima - molekularnom, ćelijskom, organizmu, populaciji i biocenotičkom. Reparativni procesi idu uz trošenje energije i plastičnih tvari, stoga je reparacija moguća samo ako se održava dovoljna brzina metabolizma. Ako se metabolizam zaustavi, prestaje i reparacija. U ekstremnim uslovima spoljašnje sredine očuvanje disanja je posebno važno, jer upravo disanje obezbeđuje energiju za procese reparacije.
Redukciona sposobnost ćelija adaptiranih organizama određena je otpornošću njihovih proteina na denaturaciju, odnosno stabilnošću veza koje određuju sekundarnu, tercijarnu i kvartarnu strukturu proteina. Na primjer, otpornost zrelog sjemena na visoke temperature obično se povezuje s činjenicom da nakon dehidracije njihovi proteini postaju otporni na denaturaciju.
Glavni izvor energetskog materijala kao supstrata za disanje je fotosinteza, stoga opskrba energijom ćelije i povezani procesi reparacije zavise od stabilnosti i sposobnosti fotosintetskog aparata da se oporavi od oštećenja. Za održavanje fotosinteze u ekstremnim uvjetima u biljkama, aktivira se sinteza komponenti tilakoidne membrane, inhibira se oksidacija lipida, a ultrastruktura plastida se obnavlja.
Na nivou organizma, primjer regeneracije je razvoj zamjenskih izdanaka, buđenje uspavanih pupoljaka kada su tačke rasta oštećene.
Ako pronađete grešku, označite dio teksta i kliknite Ctrl+Enter.
(prema N. Green et al., 1993.)
| Adaptacija | Primjeri | ||
| Smanjenje gubitka vode | |||
| Listovi pretvoreni u iglice ili bodlje Potopljeni stomati Listovi smotani u cilindar Debela voštana kutikula Debela stabljika sa velikim odnosom zapremine i površine Dlakasti listovi Opadanje lišća tokom suše Stomati su otvoreni noću, a zatvoreni tokom dana kiseline Izdužena Henleova petlja u bubrezi Tkiva su otporna na toplotu zbog smanjenog znojenja ili transpiracije Životinje se skrivaju u jazbinama Rupe za disanje su prekrivene zaliscima | Cactaceae, Euphorbiaceae (spurge), četinari Pinus, Ammophila Ammophila Listovi većine kserofita, insekti Cactaceae, Euphorbiaceae ("sukulenti") Mnoge alpske biljke Fouguieria splendens Crassulaceae (crassulaceae) i neke biljke Crassulaceae (crassula)-4 may republic. Pustinjski sisari, npr. kamila, pustinjski pacov Mnogo pustinjskih biljaka, deva Mnogo malih pustinjskih sisara, npr. pustinjski pacov Mnogi insekti | ||
| Povećajte apsorpciju vode | |||
| Ekstenzivni plitki korijenski sistem i duboko penetrirajuće korijenje Dugo korijenje Kopanje prolaza do vode | Neke Cactaceae kao što su Opuntia i Euphorbiaceae Mnoge alpske biljke kao što su rumun (Leontopodium alpinum) Termiti | ||
| skladište vode | |||
| U mukoznim stanicama iu ćelijskim zidovima U specijalizovanoj bešici U obliku masti (voda je produkt oksidacije) | Cactaceae i Euphorbiaceae Pustinjska žaba Pustinjski štakor | ||
| Fiziološka otpornost na gubitak vode | |||
| Vidljiva dehidracija ostaje održiva Gubitak značajnog dijela tjelesne težine i brzi oporavak s dostupnom vodom | Neke epifitske paprati i mahovine, mnoge briofite i lišajevi, šaš Carex physoides Lumbricus terrestris (gubi do 70% mase), kamila (gubi i do 30%) | ||
Kraj stola 4.9
Kombinovano djelovanje temperature
I vlažnost
Uzimanje u obzir pojedinačnih faktora životne sredine nije krajnji cilj ekoloških istraživanja, već način pristupa složenim ekološkim problemima, dati komparativnu ocjenu značaja različitih faktora koji djeluju zajedno u stvarnim ekosistemima.
Temperatura i vlažnost su vodeći klimatski faktori i usko su međusobno povezani (slika 4.19).
Rice. 4.19. Utjecaj temperature na relativnu vlažnost
vazduh (prema B. Nebelu, 1993.)
Uz konstantnu količinu vode u zraku, relativna vlažnost raste kako temperatura pada. Ako se vazduh ohladi ispod tačke zasićenja vodom (100%), dolazi do kondenzacije i padavina. Kada se zagrije, njegova relativna vlažnost opada. Kombinacija temperature i vlage često igra odlučujuću ulogu u distribuciji vegetacije i životinja. Interakcija temperature i vlažnosti zavisi ne samo od relativne, već i od njihove apsolutne vrednosti. Na primjer, temperatura ima izraženiji učinak na organizme u uvjetima vlažnosti blizu kritične, odnosno ako je vlažnost vrlo visoka ili vrlo niska. Vlažnost također igra važniju ulogu na temperaturama blizu ekstrema. Dakle, iste vrste organizama u različitim geografskim područjima preferiraju različita staništa. Da, do pred pravilo, koji je ustanovio V. V. Aljehin (1951) za vegetaciju, rasprostranjene vrste na jugu rastu na sjevernim padinama, a na sjeveru se nalaze samo na južnim (sl. 4.20).
Rice. 4.20. Šema pravila anticipacije (prema V.V. Aljehinu, 1951):
1 - sjeverne vrste koje žive na uzvisini, na jugu prelaze do padina sjeverne ekspozicije i u grede; 2 - južna vrsta, nalazi se na sjeveru na najtoplijim padinama južne ekspozicije
Principi otkriveni za životinje promjena staništa(G. Ya. Bei-Bienko, 1961) i princip promjene nivoa(M. S. Gilyarov, 1970), gdje se mezofilne vrste nalaze u središtu areala, na sjeveru biraju suša mjesta, a na jugu - vlažnija mjesta ili prelaze iz kopnenog načina života u podzemni, poput mnogih insekata fitofaga. Što se slabiji uticaj klime manifestuje u specifičnim staništima koja vrsta bira, to je veća njihova sposobnost da žive u različitim klimatskim uslovima. Vrsta bira kombinaciju čimbenika koja je najprikladnija za njenu ekološku valenciju mijenjajući svoje stanište i na taj način prevazilazi klimatske granice.
Odnos između temperature i vlažnosti dobro se odražava u klimatskim dijagramima sastavljenim prema Walther-Gossen metoda, na kojoj se u određenoj skali upoređuje godišnji tok temperature vazduha sa tokom padavina (slika 4.21).

Rice. 4.21. Klimatski dijagram prema Walter-Gossenu za Odesu
(prema G. Walteru, 1968):
a - visina iznad nivoa mora; b - broj godina posmatranja temperature (prva cifra) i padavina (druga cifra); c - prosječna godišnja temperatura; d - prosječna godišnja količina padavina u mm d - prosječni dnevni minimum najhladnijeg mjeseca; e je apsolutni minimum; w - prosječni dnevni maksimum najtoplijeg mjeseca; h - apsolutni maksimum; i - kriva srednjih mjesečnih temperatura; k - kriva prosječne mjesečne količine padavina (odnos 10°=20 mm); l - isto (odnos 10°=30 mm); m - sušni period; n - polusušni period; o - vlažna sezona; n - mjeseci sa srednjom dnevnom minimalnom temperaturom ispod 0°C; p - mjeseci sa apsolutnom minimalnom temperaturom ispod 0°C, c - period bez mraza. Na apscisi - mjeseci
Klimatski dijagrami se mogu graditi za pojedine godine, a slaganjem uzastopno i kontinuirano jedan za drugim, možete dobiti klimatogram. Ekstremno sušne ili ekstremno hladne godine lako se prate na klimatogramima, što je vrlo korisno za određivanje prikladnosti kombinacija temperature i vlage u područjima gdje se očekuje unošenje biljaka ili divljači.
Atmosfera
Kao što je ranije navedeno, naša planeta Zemlja se razlikuje od drugih planeta po prisutnosti zračne ljuske, atmosfere, atmosferskog zraka. Atmosferski vazduh je mešavina raznih gasova. Sadrži 78,08% azota, 20,9% kiseonika, 0,93% argona, 0,03% ugljen-dioksida, ostalih gasova (helijum, metan, neon, ksenon, radon itd.) oko 0,01%.
Značaj atmosferskog zraka za žive organizme je ogroman i raznolik. Izvor je kisika za disanje i ugljičnog dioksida za fotosintezu. Štiti žive organizme od štetnog kosmičkog zračenja, pomaže u očuvanju topline na Zemlji.
Atmosfera je važan dio ekosfere, s kojom je povezana biogeohemijskim ciklusima, uključujući i plinovite komponente. To su ciklusi ugljenika, azota, kiseonika i vode. Fizička svojstva atmosfere takođe su od velike važnosti. Dakle, zrak pruža samo mali otpor kretanju i ne može poslužiti kao oslonac kopnenim organizmima, što je direktno utjecalo na njihovu strukturu. U isto vrijeme, neke grupe životinja počele su koristiti let kao prijevozno sredstvo. Posebno treba napomenuti da u atmosferi postoji stalna cirkulacija vazdušnih masa čiju energiju daje Sunce (slika 4.22).

Rice. 4.22. Pojednostavljena shema opće cirkulacije
vazdušne mase atmosfere:
1 - topli vazduh; 2 - hlađeni vazduh; 3 - zone visokog pritiska; CE - pasati; SD - dominantni jugozapadni vjetar; GH - polarni sjeveroistočni vjetrovi
Rezultat cirkulacije je preraspodjela vodene pare, jer ih atmosfera hvata na jednom mjestu (gdje voda isparava), prenosi i odaje na drugo mjesto (gdje padaju padavine). Ako gasovi uđu u atmosferu, uključujući zagađivače, kao što je sumpor-dioksid u industrijskim područjima, onda će ih atmosferski cirkulacijski sistem preraspodijeliti i oni će ispasti na drugim mjestima, rastvoreni u kišnici (slika 4.23).
Vjetar, u interakciji sa drugim faktorima životne sredine, može uticati na razvoj vegetacije, prvenstveno drveća koje raste na otvorenim površinama. To obično rezultira usporavanjem rasta i naginjanjem prema vjetru.
Vjetar igra važnu ulogu u širenju spora, sjemena i sl., proširujući mogućnosti za širenje nepokretnih organizama - biljaka, gljivica i nekih bakterija. Vjetar također može utjecati na migraciju letećih životinja.

Rice. 4.23. Hidrološki ciklus i skladištenje vode
(prema E. A. Kriksunov et al., 1995.)
Još jedna karakteristika atmosfere je njen pritisak, koji opada sa visinom. Evolucija živih organizama na našoj planeti odvijala se pri atmosferskom pritisku od 760 mm Hg na nivou mora i smatra se "normalnim". S povećanjem nadmorske visine, na primjer, kada se ljudi penju na planine, može doći do stanja zbog nedovoljne zasićenosti krvi kisikom. hipoksija ili anoksija. Nastaje zbog činjenice da s povećanjem nadmorske visine, parcijalni tlak kisika, kao i drugih plinova sadržanih u atmosferskom zraku, pada. Na nadmorskoj visini od 5450 m atmosferski pritisak je upola manji od nivoa mora. I iako ovdje zrak sadrži isti postotak kisika, njegova koncentracija po jedinici volumena je upola manja.
U biljkama u ovim uvjetima povećava se transpiracija, što je zahtijevalo razvoj adaptacija za očuvanje vode, kao na primjer kod mnogih alpskih biljaka.
Topografija
Topografija(reljef) se odnosi na orografske faktore i usko je povezan sa drugim abiotičkim faktorima, iako ne spadaju u takve direktne faktore sredine kao što su svetlost, toplota, voda i tlo. Glavni topografski (orografski) faktor je visina. Sa nadmorskom visinom se smanjuju prosječne temperature, povećava dnevna temperaturna razlika, povećava se količina padavina, brzina vjetra i intenzitet zračenja, smanjuje se atmosferski tlak i koncentracija plina. Dakle, povećanje nivoa terena na svakih 100 m je praćeno smanjenjem temperature zraka za oko 0,6 °C.
Ovisno o veličini oblika, topografija ili reljef se dijeli na nekoliko redova: makroreljef(planine, međuplaninske depresije, nizije), mesoreljef(brda, jaruge, grebeni, vrtače, stepski "tanjiri" itd.) i mikroreljef(mala udubljenja, neravnine, uzvisine uz stabljike, itd.), sve to utiče na biljke i životinje. Kao rezultat toga, vertikalno zoniranje je postalo uobičajeno (Slika 4.24).

Rice. 4.24. Dijagram koji pokazuje korespondenciju između uzastopnih
vertikalne i horizontalne vegetacijske zone:
1 - tropska, zona (zona tropskih šuma); 2 - umjereni pojas (zona listopadnih i četinarskih šuma); 3 - alpska zona (zona zeljaste vegetacije, mahovina i lišajeva); 4 - polarna zona (zona snijega i leda)
Planinski lanci mogu poslužiti kao klimatske barijere. Vlažan zrak se hladi dok se diže iznad planina, uzrokujući velike količine padavina na vjetrovitim padinama.
Na zavjetrinoj strani planinskog lanca formira se takozvana „kišna sjenka“, ovdje je zrak suvlji, pada manje padavina, stvaraju se pustinjski uvjeti, jer se zrak, spuštajući se, zagrijava i upija vlagu iz tla.
Utiče na žive organizme. Za većinu kralježnjaka gornja granica života je oko 6,0 km. Smanjenje tlaka s visinom podrazumijeva smanjenje opskrbe kisikom i dehidraciju životinja zbog povećanja brzine disanja. Nešto otporniji su člankonošci (prolećari, grinje, pauci) koji se mogu naći na glečerima iznad granice vegetacije. Alpske biljke karakterizira čučanj rast. U svim visokoplaninskim predjelima zemaljske kugle preovlađuju nisko rastuće puzavice i šiblje (sl. 4.25), jastučaste i rozetaste višegodišnje trave, busen i šaš, mahovine i lišajevi.

Rice. 4.25. Juniper Turkestan - na padinama grebena
Terekei-Alatau (prema I. G. Serebryakovu, 1955):
A - drvolika (livadsko-šumski pojas, 2900 mnv); B - vilenjak (subalpski pojas, 3200 m nadmorske visine)
Karakteristična morfološka osobina mnogih alpskih zdepastih biljaka, kao što su grmovi i patuljasti grmovi, je značajna prevlast podzemne mase u odnosu na nadzemnu.
Nizak rast alpskih biljaka povezan je sa adaptacijom na niske temperature i sa efektom oblikovanja zračenja, koje je bogato kratkotalasnim dijelom spektra, koji inhibira procese rasta. U anatomskoj građi alpskih biljaka postoji niz karakteristika koje doprinose zaštiti od viška zračenja, povezane su s prirodom vodnog režima i metabolizmom u visoravni: zadebljanje pokrovnog tkiva koje daje otpornost na jake vjetrove itd. Biljke koje žive na stijenama doživljavaju promjene u pravcu kseromorfoze: smanjuju se veličine ćelija i povećava gustina tkiva, povećava se broj zubaca po jedinici površine lista, smanjuju se njihove veličine. Kod vrsta koje žive u blizini otopljene vode ili drugih izvora vlage, listovi su veći, a kseromorfne karakteristike su manje izražene.
Niske temperature i jaka svjetlost pospješuju stvaranje velikih količina antocijana, pa stoga duboki, bogati tonovi boje cvijeća. Kombinacija malih listova s malim rastom i velikih cvjetova jarkih boja karakteristična je karakteristika mnogih alpskih biljaka.
Karakteristična karakteristika fiziologije i biohemije visokoplaninskih biljaka je povećanje intenziteta redoks procesa, povećanje aktivnosti enzima koji su u njima uključeni (katalaza, peroksidaza itd.) i niži temperaturni optimum za njihov rad. nego u nizinskim biljkama.
Disanje alpskih biljaka otporno je na štetne učinke, u pravilu se uočava povećanje disanja, a samim tim i povećanje energije koja se oslobađa pri razgradnji složenih spojeva. Prema savremenim konceptima, ovo je jedan od fizioloških temelja adaptacije biljaka na ekstremne uslove.
Prilikom penjanja u planine mijenja se i sezonski razvoj biljaka. Dakle, u proleće, penjući se na planine, može se videti razvoj iste vrste u sledećem redosledu: u niskoplaninskoj zoni - cvetanje, u proseku - pupanje, čak i više - početak vegetacije, i, konačno, samo izgled nakon što se snijeg otopi. U jesen, prilikom penjanja na planine, uočavamo ubrzani početak jesenjih fenofaza: bojenje lišća, opadanje listova i odumiranje nadzemnih dijelova. Jasno je smanjenje vegetacijskog perioda kod biljaka.
Uz nadmorsku visinu, ekspozicija i strmina padina su od velikog značaja za žive organizme.
Na sjevernoj hemisferi, planinske padine okrenute prema jugu dobijaju više sunčeve svjetlosti, intenzitet svjetlosti i temperatura su ovdje viši nego na dnu doline i na padinama sjeverne ekspozicije. Na južnoj hemisferi situacija je obrnuta. Ovo ima upečatljiv učinak i na prirodnu vegetaciju i na zemljište koje koristi čovjek. Na primer, široke pukotine između stena iznad Dunava u istočnoj Srbiji, zaštićene od vetrova i koje doživljavaju vlažni efekat reke, doprinele su očuvanju mnogih retkih, reliktnih i endemičnih biljnih vrsta, među kojima je i „medveđa leska“ - Corylus. colurna, orah - Juglans regia, jorgovan (divlji oblik) - Syringa vulgaris i dr.
Strme padine karakteriziraju brza drenaža i erozija tla. Ovdje su tla obično tanka i suva, sa kseromorfnom vegetacijom. Sa nagibom većim od 35 °, tlo se ne formira, nema vegetacije, stvoreni su seluci od rastresitog materijala.
Ostali fizički faktori
Ostali fizički faktori koji okružuju žive organizme na Zemlji uključuju uglavnom atmosferski elektricitet, vatra, buka, Zemljino magnetno polje, jonizujuće zračenje.
atmosferski elektricitet djeluje na žive organizme putem pražnjenja i jonizacije zraka. Na primjer, poznat je destruktivni učinak groma kada udari u velika stabla i životinje. Postoje određeni obrasci u učestalosti oštećenja od groma na raznim vrstama drveća. To je povezano i s oblikom krošnje i s električno vodljivim svojstvima kore, na primjer, sa brzinom njenog vlaženja. Po učestalosti udara groma na prvom mjestu su smreka i bor, zatim breza, a jasika se znatno rjeđe oštećuje. Grom uzrokuje mehanička oštećenja drveća (pocijepana debla, pukotine), gubitak krupnih stabala, čime utiče na strukturu šumske sastojine, često izazivajući požare. Oko 21% šumskih požara u Rusiji nastaje zbog udara groma, tokom grmljavine.
Uloga atmosferskih električnih pražnjenja je i u tome što za vrijeme grmljavine sintetiziraju dušikove okside iz atmosferskog dušika i kisika, koji s kišnicom ulaze u tlo i akumuliraju se u njemu od 4 do 10 kg godišnje po 1 hektaru u obliku nitrata. i azotne kiseline.
Učinak ionizacije zraka na ljude, životinje i biljke još nije dovoljno proučavan. Istovremeno, pouzdano je utvrđena direktna veza između blagostanja osobe i prisustva lakih jona u zraku. Izraženo je mišljenje da ionizacija zraka služi materijalnoj sposobnosti nekih biljaka da „predviđaju vrijeme“ (smanjenje fotosinteze i disanja, zatvaranje stomata i prestanak transpiracije prije grmljavine mnogo prije pada atmosferskog tlaka). Eksperimentalno je dokazano djelovanje slabe struje na korijenski sistem nekih biljaka. Na primjer, u sadnicama smreke i bora fitomasa se povećava za 100-120%. Utvrđeno je da je djelovanjem usmjerenog električnog polja moguće regulisati brzinu kretanja tvari unutar stabla, a samim tim i brzinu njegovog rasta.
Vatra u životu biljaka i životinja - prilično rijedak, ali vrlo učinkovit faktor. Požari, na primjer, u šumama, kao što je ranije navedeno, mogu nastati i prirodno od udara groma i krivnjom čovjeka, njegovih aktivnosti. Stoga se vatra pripisuje i prirodnim okolišnim faktorima i antropogenim.
Ozbiljne posledice nisu samo krunisani šumski požari, koji zahvataju čitavu šumsku sastojinu, već i podzemne površine, koji uništavaju prizemnu vegetaciju, šikaru, niže grane drveća, a često i korenov sistem. Životinje umiru. Osim štete direktno od požara, požari uzrokuju propadanje šumske sastojine. Rast se smanjuje. Oslabljena stabla su podložnija gljivama, poput truleži drveta, koje lako prodiru kroz „vatrene rane“, a napadaju ih štetnici.
Šumski požari uvelike mijenjaju uslove života biljaka i životinja. Tokom požara u četinarskim šumama temperatura dostiže 800-900°C, u zemljištu na dubini od 3,5 cm - do 95°C, na dubini od 7 cm - do 70 "C. U suvim šumama, stelja i humus u tlu skoro potpuno sagorijeva Mineralne čestice gornjeg sloja tla se sinteruju Nastaju grudice ili staklasta kora koja teško prožima zrak, vodu i korijenje Zemljište je jako zbijeno Zbog sagorijevanja organskih kiselina i oslobađanjem baza, kiselost tla naglo opada, u gornjim horizontima pH vrijednost često dostiže jako alkalnu Od visoke temperature gornji slojevi tla se steriliziraju - mikroflora tla umire, au dubljim slojevima se mijenja njen sastav, grupe koje su najvažnije za život biljaka su osiromašene. Tako u zemljištima četinarskih šuma nakon požara prevladava aktivnost mikroorganizama koji izazivaju maslačnu fermentaciju i denitrifikaciju.
Nakon šumskih požara dolazi do nagle promjene uslova u biljnim zajednicama (rasvjetljavanje, promjena temperature i drugih mikroklimatskih faktora), posebno kada je šumska sastojina uništena, što dovodi do toga da se u budućnosti opožarena područja naseljavaju vrstama. živih organizama sa različitim adaptivnim osobinama koje pomažu da izdrže vatru i prežive.na brdima. Dakle, kod biljaka su to duboki podzemni pupoljci obnavljanja, sposobnost sjemena da dugo ostane u tlu i izdrži visoke temperature, izdržljivost na mraz, jako svjetlo itd.
Revegetacija na izgorjelim područjima ima svoje karakteristike. Pionirske mahovine pojavljuju se na spaljenim mjestima od spora koje raznosi vjetar; nakon tri do pet godina, "vatrena mahovina" - Funaria hygrometrica - je najzastupljenija mahovina. Od viših biljaka, Ivan-chai (Chamaenerion angustifolion) brzo naseljava vatru. Postepeno naseljavanje opožarenih površina dešava se i sa drvenastim rastinjem - vrba, breza, jasika itd. (Sl. 4.26).

Rice. 4.26. Utjecaj vatre na vegetaciju "klinova" drveća
Trans-Uralska šumska stepa (prema D. F. Fedyunin, 1953):
A - prije požara; B - nakon požara; B - godinu dana nakon požara; 1 - vrba; 2 - breza, 3 - jasika
Stepski požari („opekotine“) mogu biti manje-više redoviti, povezani s ljudskim aktivnostima, i igraju bitnu ulogu u životu živih organizama, ponekad pozitivnu na regulaciju rasta, obnavljanje, odabir vrsta i održavanje stalnog sastava bilja.
Buka kao prirodni faktor životne sredine za žive organizme je beznačajan, ali može imati i značajan uticaj sa povećanim antropogenim uticajima (buka nastala tokom rada vozila, opreme industrijskih i kućnih preduzeća, ventilacionih i gasnoturbinskih instalacija itd.) .
Vrijednost zvučnog pritiska se mijenja i normalizira u decibelima. Cijeli raspon zvukova koji ljudi čuju je unutar 150 dB. Na našoj planeti život organizama odvija se u svijetu zvukova. Na primjer, ljudski organ sluha je prilagođen određenim stalnim ili ponavljajućim zvukovima (auditivna adaptacija). Osoba gubi sposobnost za rad bez uobičajene buke. Glasna buka je još štetnija po ljudsko zdravlje. Ljudi koji žive i rade u nepovoljnim akustičnim uslovima pokazuju znakove promjena u funkcionalnom stanju centralnog nervnog i kardiovaskularnog sistema.
Istraživanja su dokazala uticaj buke na biljne organizme. Tako biljke u blizini aerodroma, sa kojih kontinuirano polaze mlazni avioni, doživljavaju inhibiciju rasta, pa čak i nestanak pojedinih vrsta. Niz naučnih radova pokazao je depresivno dejstvo buke (oko 100 dB sa frekvencijom zvuka od 31,5 do 90 hiljada Hz) na biljke duvana, gde je uočeno smanjenje intenziteta rasta listova, prvenstveno kod mladih biljaka. Pažnju naučnika privlači i efekat ritmičkih zvukova na biljke. Istraživanja o uticaju muzike na biljke (kukuruz, bundeva, petunija, cinija, neven), koju je 1969. godine sproveo američki muzičar i pevač D. Retolak, pokazala su da biljke pozitivno reaguju na Bahovu muziku i indijske muzičke melodije. Njihov habitus, suha masa biomase bila je najveća u poređenju sa kontrolom. I ono što je najviše iznenađujuće je da su se njihove stabljike zaista pružale prema izvoru ovih zvukova. Istovremeno, zelene biljke su na rok muziku i neprekidne ritmove bubnjeva reagovale smanjenjem veličine listova i korijena, smanjenjem mase, a sve su se udaljile od izvora zvuka, kao da žele pobjeći od destruktivnog. efekat muzike (slika 4.27).

Rice. 4.27. Vrste biljaka nakon djelovanja različite muzike:
A - Indijske melodije (R. Shankar); B - muzika J.-S. Bach; B - rok muzika (eksperimenti D. Retolaka, 1969)
Biljke, kao i ljudi, reaguju na muziku kao integralni živi organizam. Njihovi osjetljivi "nervni" provodnici, prema brojnim naučnicima, su snopovi floema, meristemi i ekscitabilne ćelije smještene u različitim dijelovima biljke, međusobno povezane bioelektričnim procesima. Vjerovatno je ta činjenica jedan od razloga sličnosti reakcije na muziku kod biljaka, životinja i ljudi.
Zemljino magnetsko polje. Naša planeta Zemlja ima magnetna svojstva. Igla kompasa je uvijek orijentirana duž magnetnog meridijana, usmjerena na sjever na jednom kraju i na jug na drugom. Magnetolozi su pokazali da je za stvaranje vidljivog geomagnetskog polja u centru Zemlje potrebno postaviti džinovski cilindrični magnet prečnika 200 km i dužine 4000 km. Osa zemljinog magneta nalazi se pod uglom od 1,5" u odnosu na osu rotacije Zemlje, tako da se magnetni polovi ne poklapaju sa geografskim. Vremenom magnetni polovi menjaju svoj položaj. Utvrđeno je da sjeverni magnetni pol pomiče se iznad površine Zemlje za 20,5 m dnevno, odnosno 7,5 km godišnje, a južni - za 30 m (11 km godišnje). Kao i svaki magnet, Zemljine magnetne linije sile izlaze iz jedan pol i zatvaraju se kroz prostor blizu Zemlje u drugom polu. Zbog ovog fenomena nastaje magnetosfera u blizini Zemlje (slika 4.28).

Rice. 4.28. Meridijalni presjeci Zemljine magnetosfere:
1 - solarni vetar; 2 - prednji udar; 3 - magnetna šupljina; 4 - magnetopauza; 5 - gornja granica magnetosferskog jaza; 6 - plazma plašt; 7 - vanjski pojas za zračenje; 8 - unutrašnji radijacioni pojas, ili plazmasfera; 9 - neutralni sloj; 10 - plazma sloj
Odgađa protok solarnih nabijenih čestica, zvanih plazma, ili solarni vjetar, sprječavajući ih da stignu do površine planete. Sunčev vjetar, takoreći, obilazi Zemlju i pomiče se na noćnu stranu, povlačeći, zauzvrat, magnetne linije sile u istom smjeru. Deformacija linija magnetnog polja nastaje zbog činjenice da tokovi solarne plazme nose sa sobom, takoreći, "zamrznuto" magnetno polje, koje je u interakciji sa Zemljinom magnetosferom. Tokom proteklih 600 hiljada godina, paleomagnetolozi su zabilježili 12 epoha preokreta geomagnetskog polja (tabela 4.10).