Акцептором со2 в с4 пути фотосинтеза является. Сравнительная характеристика -С3 и -С4 растений. Липиды: классификация и их роль в растении
Альтернативные пути фотосинтеза.
С 3- путь оказался не единственным в превращении Со 2 а превращении в темновой фазе. Изменение внешних условий климата и состава ат мосферы в разных участках Земли, вызвали появление континентов с жарким и засушливым климатом (пустыни), что не могло не сказаться на растительном мире. Видовая специфичность несомненно является отражением факта своеобразия физиологического типа каждого вида растения. C 50 годов обратили внимание на процессы фотосинтеза, дыхания, транспирации, транспорта веществ. Оказалось, что условия среды определяют структурно-биохимические типы фотосинтеза растений.
Вначале 60 годов 20 века было установлено, что у некоторых растений произрастающих в тропиках или вышедших оттуда, как кукуруза, сахарный тростник, сорго, просо, представители семейства маревых растений полупустыни и пустыни, всего у 18 семейств. Такие растения встречаются как различных семействах, так и родах, но нет ни одного семейства, рода, состоящих только из С 4- растений. Первые продукты ассимиляции СО 2 здесь имеют 4 атома углерода, поэтому такие растения называют "С4-растениями". У этих растений стабильным первичным продуктом ассимилируемой СО 2, является не ФГК, а яблочная, а у ряда растений аспарагиновая кислота, содержащих не 3 , а 4 углеродных атома., в которых была обнаружена радиоактивная метка 14 СО 2 (Ю.С. Карпилов, 1960,1969; М.Д. Хэтч и С.Р. Слэк, 1966). Так был открыт по существу новый путь ассимиляции СО 2, которой получил название «С 4 – пути» фотосинтеза.
У таких растений специфичное анатомическое строение листа. У них два типа фотосинтезирующих клеток: клетки обкладки, радиально покрывающие проводящие пучки в листе, и клетки мезофилла столбчатой и губчатой ткани. Это может служить диагностическим признаком на определение С 4 -растений. В структуре этих клеток также есть различия: в клетках обкладках в хлоропластах отсутствуют граны, они агранальные, а в клетках мезофилла все хлоропласты гранальные. Эти два типа клеток и физиологически не равноценны и специализируются на выполнении различных звеньев в превращении поглощенного СО 2
Клетки обкладки выполняют основную функцию, в них происходит цикл Кальвина, в том виде как он происходит у растений с С 3 –путем фотосинтеза, т.е. весь процесс превращения углекислого газа всех этапов цикла Кальвина. Клетки мезофилла листа осуществляют вспомогательную роль, подкачку СО 2 для цикла Кальвина. Таким образом у растений с С 4 –путем. имеется две стадии, происходящих в разных клетках листа. На один агранальный хлоропласт клетки обкладки приходится 10 гранальных клеток мезофилла, в которых происходит нециклический путь фотофосфорилирования. Это обеспечивает повышение фотосинтеза даже в условиях очень высокой температуры и почти закрытых устьицах, что имеет положительное значение и для водного режима растений, т.к снижается расход воды. Оба процесса пространственно разобщены, т.е усвоение СО 2 идет в клетках мезофилла, а образование сахаров и регенерация акцептора в клетках обкладках. Но эти два процесса идут одновременно, на свету.
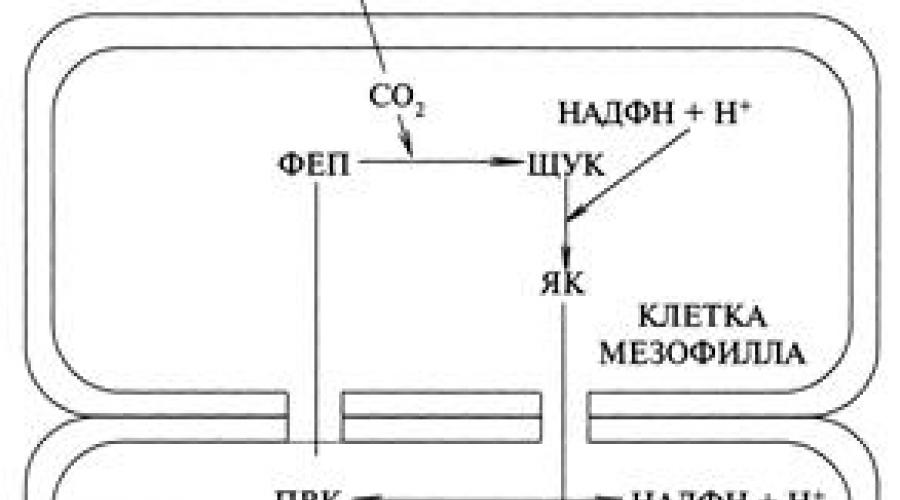
Первая реакция цикл С 4 - пути фотосинтеза протекает в мезофилле листа, в котором акцептором углекислого газа является фосфоэнол-пировиноградная кислота (ФЭП). В этих клетках очень активна ФЭП-карбоксилаза, интенсивно связывающая СО 2 в органическое соединение (рис.)

Фотосинтетический цикл С 4 –пути фотосинтеза
ФЕП - фосфоенолпировиноградная кислота;
ЩУК - щавелевоуксусная кислота;
ЯК - яблочная кислота;
ПВК - пировиноградная кислота;
РБФ - рибулозо-1,5-бифосфат;
3-ФГК - фосфоглицериновая кислота;
ФГА - фосфоглицериновый альдегид
Реакции идут следующим путем. В клетках мезофилла листа происходит карбоксилирование ФЭП ферментов ФЭП-карбоксилаза с образованием ЩУК щавелево-уксусной кислоты: ФЭП + СО 2 + Н 2 О → ЩУК Это нестабильное вещество стабилизируется здесь же за счет НАДФН с образованием яблочной кислоты (малат), 4-х углеродного соединения: ЩУК + НАДФН → ЯК. У ряда растений вместо ЯК, образуется аспарагиновая кислота (АК) для чего в клетках необходим аммиак: ЩУК + NH 4 + НАДФН → АК. Эти две кислоты в одном и том же растении образовываться не могут, поэтому такие растения называю малатными или аспартатными.. Второй этап заключается в том, что малат переходит в клетки обкладки сосудистого пучка, где в результате его окислительного декарбоксилирования образуются СО 2 и НАДФН для цикла Кальвина. Получаемый при этом пируват (ПВК) возвращается в клетки мезофилла фосфорилируется, в фосфоенолпируват, первичный акцептор СО 2, и С4-путь замыкается. Освободивщаяся СО 2 идет на второе карбоксилирование уже РДФ цикла Кальвина с образованием углеводов.в клетах обкладках. Двойное карбоксилирование обеспечивает усвоение CO 2 более интенсивно, чем при С 3 – пути, поэтому его называют кооперативным. Двоное карбоксиливание намного эффективнее и обеспечивает интенсивный фотосинтез при пониженном содержании СО 2 . Так если у С 3 растений фотосинтез снижается ниже 0,03%, то у С 4 – идет полноценно при 0,008%. Появление альтернативного пути С 4 растений считают как процесс эволюции приспособления с пониженному содержания в атмосфере СО 2 и увеличения О 2 , за с чет самих же растений, чем в период их возникновения. Поэтому у этих растений большие преимущества перед С 3 – путем.
Путь С4 получил свое название, т.к. в темновой фазе первичным продуктом фиксации СО2 в этом случае является органическое соединение не с тремя, а с четырьмя атомами углерода (щавелевоуксусная кислота). Таким типом фотосинтеза обладают тропические растения жарких стран, например, бромелиевые. Было давно замечено, что эти растения усваивают СО2 намного лучше С3 растений. В анатомической структуре листьев С4-растений, наряду с нормальными обычными хлоропластами, вокруг сосудистых пучков у них имеются особый вид очень плотных хлоропластов почти без тилакоидов, но наполненных крахмалом. Эти хлоропласты назвали обкладочными.
В обычных хлоропластах у С4 растений, как и положено, протекает световая фаза фотосинтеза, а также происходит фиксация СО2 , но при этом образуется щавелевоуксусная кислота. Такая щавелевоуксусная кислота превращается в яблочную, которая поступает в обкладочные хлоропласты, где сразу расщепляется с выделением СО2. А дальше все идет, как и у нормальных С3 растений. При этом концентрация СО2 в обкладочных хлоропластах в результате становится значительно выше, чем у С3 растений, а очень плотное расположение этих хлоропластов обеспечивает то, что кислорода к ним почти не поступает, межклетников-то нет. Поэтому, раз нет кислорода, а углекислого газа сколько хочешь, фотодыхание не наступает.
Таким образом, у С4 растений и фиксация СО2 происходит более эффективно в виде других соединений, и образование сахаров осуществляется в особых хлоропластах, в результате чего сокращается интенсивность фотодыхания и связанных с ним потерь.
С4 растения могут захлопнуть свои устьица в жару, и не терять такой драгоценной влаги. У них обычно накоплено достаточно СО2 в виде яблочной кислоты.
27. Фотодыхание: биохимические реакции, их локализация. Физиологическая роль фотодыхания.
Фотодыхание – это активируемый светом процесс выделения СО2 и поглощения О2. Первичным продуктом фотодыхания является гликолевая кислота. Фотодыхание усиливается при низком содержании СО2 и высокой концентрации О2 в воздухе. В этих условиях рибулозодисфаткарбоксилаза хлоропластов катализирует не карбоксилирование рибулозо-1,5-дифосфата, а его расщепление на 3-фосфоглицериновую и 2-фосфогликолевую кислоты. Последняя дефосфорилируется с образованием гликолевой кислоты.
Гликолевая кислота из хлоропласта переходит в пероксисому, где окисляется до глиоксиловой кислоты. Глиоксиловая кислота аминируется, превращаясь в глицин. Глицин транспортируется в митохондрию, где из двух молекул глицина синтезируется серин и освобождается СО2.
Серин может поступать в пероксисому и передает аминогруппу на пировиноградную кислоту с образованием аланина, а сам превращается в гидроксипировиноградную кислоту. Последняя при участии НАДФН восстанавливается в глицериновую кислоту. Она переходит в хлоропласты, где включается в цикл Кальвина
У растений С4 -типа выделяющийся при фотодыхании углекислый газ реагирует в клетках мезофилла с фосфоэнолпировиноградной кислотой с образованием щавелевоуксусной и яблочной кислот. Яблочная кислота переходит в клетки обкладки, где служит донором СО2. Растения С3-пути характеризуются высокой интенсивностью фотодыхания. Фосфогликолевая кислота через ряд превращений распадается с выделением С02. Таким образом, при фотодыхании часть промежуточных продуктов фотосинтеза теряется за счет выделения С02. Реакции окисления и карбоксилирования конкурируют друг с другом, а осуществление карбоксилазной или оксигеназной функции зависит от содержания 02 и С02
Фотодыхание снижает эффективность фотосинтеза, приводит к потерям ассимилированного углерода, однако имеет некоторое синтетическое значение. На ранних этапах развития жизни, когда в атмосфере было мало кислорода, рубиско заняло ключевую позицию в фотосинтезе, и ее оксигеназная функция не доставляла проблем. По мере увеличения содержания кислорода потери на фотодыхание нарастали, и у ряда растений возникли механизмы активной доставки к месту работы рубиско углекислого газа (см. C4 и CAM-фотосинтез), увеличивающее долю её карбоксилазной активности до 100%.
Фотосинтез – это процесс трансформации поглощенной растением энергии солнечного света в химическую энергию органических соединений. С 4 - путь фотосинтеза или цикл Хетча-Слэка.
Австралийскими учеными Хетчем и Слэком был описан С 4 -путь фотосинтеза, характерный для тропических и субтропических растений (сахарный тростник, кукуруза и др.). Листья этих растений содержат хлоропласты двух типов: обычные в клетках мезофилла и крупные хлоропласты, не имеющие гран и фотосистемы II, в клетках обкладки, окружающих проводящие пучки.
В цитоплазме клеток мезофилла присоединяет СО 2 к пировиноградной кислоте, образуя щавелевоуксусную кислоту. Она транспортируется в хлоропласты, где восстанавливается до яблочной кислоты при участии НАДФН. В присутствии ионов аммония щавелевоуксусная кислота превращается в аспарагиновую кислоту. Яблочная и (или) аспарагиновая кислоты переходят в хлоропласты клеток обкладки, восстанавливаются до пировиноградной кислоты и СО 2 . СО 2 включается в цикл Кальвина, а пировиноградная кислота переносится в клетки мезофилла, где превращается в пировиноградную кислоту. Такой механизм позволяет растениям фотосинтезировать при закрытых из-за высокой температуры устьицах. Кроме того, продукты цикла Кальвина образуются в хлоропластах клеток обкладки, окружающих проводящие пучки. Это способствует быстрому оттоку фотоассимилятов и тем самым повышает интенсивность фотосинтеза.
Липиды: классификация и их роль в растении.
Липиды представляют собой достаточно сложные по химической структуре вещества. В их состав также входят углерод, кислород, водород, но в отдельные группы липидов могут входить и фосфор, и сера, и азот (фосфатиды, пигменты). Все липиды гидрофобны, т.е. не растворяются в воде. Функции у липидов различны в зависимости от химического строения. Липиды не являются биополимерами.
Липиды классифицируются на 5 больших групп по признаку функции и сложности строения:Жиры- наиболее легко синтезируемая группа липидов. С химической точки зрения - это эфиры жирных кислот и глицерина.Основные функции жиров - энергетическая, строительная и запасающая.Воска - это жироподобные вещества, твердые при комнатной температуре. По химической структуре - это сложные эфиры между жирными кислотами и высокомолекулярными одноатомными спиртами жирного ряда.Основная функция восков - защитная.Фосфатиды - к которым относятся глицерофосфатиды, лецитины и кефалины - это молекулы сложных эфиров глицерина, жирных кислот и фосфорной кислоты. Эти вещества входят в состав запасных жиров и предохраняют их от прогоркания.Основная функция фосфатидов - запасающая.
Пигменты (хлорофиллы и каротиноиды) - это особая группа липидов, имеющая сложное строение, куда входят и азотистые радикалы. К пигментам относят две группы веществ - хлорофиллы и каротиноиды.
Основная функция пигментов - участие в энергетической (световой) фазе фотосинтеза.Стероиды - это производные сложного гетероциклического соединения. В эту группу соединений входят высокомолекулярные спирты (стеролы) и их сложные эфиры (стериды) Наиболее известный стероид - эргостерол, из которого в промышленности получают витамин Д.
Основная функция стероидов- строительная (участвуют в составе мембран).
УДК 581.1:577.1
ЭВОЛЮЦИОННЫЕ АСПЕКТЫ С4-ФОТОСИНТЕЗА
В.В. Иванищев
Рассмотрены вопросы, касающиеся многообразия проявлений фотосинтетической ассимиляции углекислого газа у растений и эволюционных аспектов С4-типа фотосинтеза. Показано, что за последние полтора десятилетия получены экспериментальные данные, которые существенным образом меняют наши представления о путях возникновения С4-фотосинтеза и реализации его особенных свойств у ряда типичных С3-растений. Обсуждаются перспективы использования таких знаний для выработки стратегии по улучшению эффективности фотосинтеза С ¡-растений для повышения их продуктивности.
Ключевые слова: растения, ассимиляция неорганического углерода, С3-С4-фотосинтез, С4-фотосинтез, эволюционные аспекты, продуктивность растений.
Введение
Фотосинтез - главный путь метаболизма, благодаря которому на нашей планете идет преобразование неорганического углерода в органические соединения. Исследование механизмов фотосинтетического процесса привело к формированию представлений о биохимических путях ассимиляции неорганического углерода С3- и С4-типов. После открытия оксигеназной реакции Рубиско ф-рибулозо-1,5-бисфосфат-карбоксилазы-оксигеназы, КФ 4.1.1.39) было принято говорить о фотодыхании, как параллельном процессе . Отдельные специалисты, в т.ч. автор, внесший значительный вклад в представления о биохимии фотодыхания, стали выделять и так называемый С2-фотосинтез . Обнаруженные особенности транспорта (не в митохондрии и пероксисомы этой же клетки) и метаболизма там фосфогликолата (образованного при окислении кислородом D-рибулозо-1,5-бисфосфата), а его перенос и метаболизм в клетках обкладки с последующим «улавливанием» выделяющегося СО2 хлоропластами в этих клетках позволили сформулировать представления о С2-фотосинтезе . В таком случае даже оксигеназная реакция Рубиско рассматривается как поставщик СО2, в результате чего общая эффективность фотосинтеза, хотя бы, не сильно снижается.
Активные исследования растений, чья жизнь приурочена к специфическим экологическим условиям, за последние три десятилетия позволили обнаружить некоторые вариации в проявлении разных путей метаболизма углерода при фотосинтезе. При этом выявлена высокая морфологическая и биохимическая пластичность как С3-, так и С4-растений. Кроме того, было открыто многообразие механизмов, за счет которых растения С3-типа проявляли ряд свойств, характерных для растений С4-типа .
В специфических экологических условиях засоления и засухи Центральной Азии были обнаружены растения, обладающие С4-подобным путем фотосинтеза без классической Кранц-анатомии листа . Более того, благодаря еще одному открытию стирается резкая грань между двумя основными типами растений по механизму фиксации СО2. Показано, что в клетках, окружающих центральную жилку проводящей системы листьев риса (типичного С3-растения), присутствуют ключевые ферменты, характерные для С4-типа фотосинтеза .
Всё это делает необходимым провести обобщение полученных результатов с целью определения дальнейших направлений исследований в данной области науки и возможных путей повышения эффективности фотосинтеза (и продуктивности) классических С3-растений, которые составляют большинство среди важнейших сельскохозяйственных культур.
Анатомо-морфологические особенности и пластичность
С4-фотосинтеза
Растения с С4-типом фотосинтеза обладают, как известно, целым комплексом особенностей. Они обеспечивают растениям ряд важных характеристик практического характера - большую устойчивость к некоторым факторам среды (повышенной температуре, засухе), более экономный расход воды, большую продуктивность при меньшем расходе азота , что объясняет экономическую заинтересованность производителей при возделывании подобных сельскохозяйственных культур. Характерные свойства структуры листа, в первую очередь, обеспечивают С4-растениям СО2-концентрирующий механизм, позволяющий предотвращать потери углекислого газа, и тем самым снижать фотодыхание, из-за которого эффективность фотосинтеза у С3-растений может уменьшаться на величину до 40%, особенно в условиях повышенной температуры и засухи .
У С3-растений можно наблюдать два физиологически и пространственно разделенных типа клеток, а именно: одна группа более или менее равномерно распределена и образует мезофилл, другая часть клеток сосредоточена вокруг проводящих пучков. Такая картина делает их похожими на С4-растения. Однако биохимически такие клетки одинаковы.
С другой стороны, некоторые растения С3-типа проявляют характеристики С4-растений в отсутствие двух анатомически существенно различных типов клеток в соответствии с классическими представлениями о С4-пути фиксации углекислого газа . Несмотря на это, С4-фотосинтез характеризуется рядом строго специфических функциональных особенностей. Понимание эволюционных преобразований в этом аспекте
базируется на представлениях об изменении характеристик клеток и их генетическом предопределении.
У большинства С4-растений функционирование ФЕП-карбоксилазы и цикла Кальвина пространственно разделено по таким двум типам клеток. Высокая плотность жилок проводящих пучков приводит к тому, что соотношение объемов тканей, образованных клетками мезофилла и обкладки сосудистых пучков, составляет примерно 1:1 .
Изучение образования этих типов клеток показывает, что в ходе онтогенеза внутренние слои клеток могут дифференцироваться из клеток прокамбия, а внешние слои - из клеток меристемы. Филогенетический анализ показывает, что С4-растения появились благодаря нескольким разным путям эволюционных изменений. При этом стенки клеток обкладки проводящих пучков могут содержать суберин, который ограничивает диффузию СО2 за пределы таких клеток. В то же время аналогичный слой суберина обнаруживают и у некоторых С3-растений на внешних стенках внутренних слоев обкладки сосудистых пучков. Таким образом, никаких характеристик, которые специфичны только для клеток листьев С4-растений, до сих пор не обнаружено .
Другим важным моментом является расстояние между клетками двух типов для обеспечения максимальной скорости обмена метаболитами между ними. Это объясняет то, что количество слоев клетки обкладки ограничивается одним-двумя. В то же время повышение эффективности процесса фотосинтеза достигается, с другой стороны, и небольшим количеством клеток мезофилла между клетками обкладки, число которых может составлять у некоторых видов до четырех. При этом количество клеток между проводящими пучками в целом - невелико. Чтобы уменьшить расстояние между двумя типами клеток, в листьях иногда встречаются бесхлорофилльные клетки .
В то же время изучение разнообразных двудольных С4-растений показало, что в сравнении с типичными С3-растениями многие их анатомо-морфологические характеристики оказались сходными. Это касается таких показателей как плотность жилок на 1 мм2 поверхности листа, толщина листа, процент межклеточного пространства. Другие показатели имели более широкие или узкие пределы изменчивости, либо они были сдвинуты в сторону более высоких или низких величин .
Третьим важным обстоятельством, характерным для С4-растний, является большой компартмент, обеспечивающий максимально возможное связывание (без потерь) выделяемого СО2 при декарбоксилировании четырехуглеродного соединения. Это объясняет наличие увеличенного количества хлоропластов, присутствующих в клетках обкладки. Превышение объема клеток обкладки над объемом клеток мезофилла достигается несколькими путями, а именно: за счет увеличения числа хлоропластов в клетках обкладки, увеличения числа клеток обкладки, либо
Количества проводящих пучков или изменения диаметра жилок . Описанные показатели являются взаимозависимыми и, отчасти, определяются экологическими условиями, которые обеспечивают, в первую очередь, необходимые параметры водного обмена и сопутствующие показатели, такие как гидравлическое давление, скорость поступления и передвижения воды в растении и т.п. Всё это обусловливает разнообразие анатомического проявления С4-синдрома.
Заключительная особенность С4-растений состоит в том, что хлоропласты клеток обкладки сосудистых пучков и клеток мезофилла различаются не только по числу, но и морфологически, и функционально. В то же время, большое количество хлоропластов в клетках обкладки характерно не только для С4-растений, но и для растений, с так называемым, С2-фотосинтезом, при котором, образуемый при фотодыхании глицин, переносится из клеток мезофилла в клетки обкладки, где, выделяемый при его метаболизме СО2, далее захватывается хлоропластами и используется в цикле Кальвина . Такие особые черты реализации этого типа фотосинтеза рассматриваются исследователями как один из путей, благодаря которому эволюция механизмов фотосинтетической ассимиляции неорганического углерода может быть представлена в виде последовательности: С3^С2^С4 . К другим характерным чертам С4-синдрома можно отнести особенности распределения между разными клетками листьев растений таких органелл как митохондрии и пероксисомы, а также особенности ультраструктуры и фотохимических свойств хлоропластов .
Описанные выше показатели весьма пластичны. В зависимости от условий среды они могут существенно меняться, благодаря чему растения с С3-путем фотосинтеза могут приобретать черты, характерные для С4-пути. В то же время многообразие возможных анатомо-морфологических изменений различных черт у С4-растений позволяет им поддерживать свои физиолого-биохимические характеристики на необходимом уровне. Однако в целом пластичность С3-растений в этом аспекте оценивается выше, несмотря на то, что экспериментальных данных в этой области явно не хватает .
Биохимические особенности проявлений С4-фотосинтеза
у высших растений
Классические общие представления о С4-пути фиксации неорганического углерода включают три основных разновидности использования первого стабильного продукта - четырехуглеродного соединения (малата и/или аспартата) в качестве источника СО2 для D-рибулозо-1,5-бисфосфат-карбоксилазы-оксигеназы (Рубиско) - ключевого фермента цикла Кальвина. Такие формы растений обозначают как НАДФ-
малик-энзимная, НАД-малик-энзимная и ФЕП-карбоксикиназная - по названию ключевого фермента, обеспечивающего декарбоксилирование четырехуглеродного соединения с выделением СО2 . Позднее у трав С4-типа были обнаружены 14 разновидностей анатомо-биохимических комбинаций . Они различались по таким характеристикам, как размеры клеток мезофилла и клеток обкладки сосудистых пучков, общие объемы клеток разного типа, особенности расположения хлоропластов в клетках, наличие числа гран в хлоропластах и т.д. с соответствующими вариациями в распределении по клеткам и величине активности ферментов.
В противоположность этим травам у двудольных известны только два типа растений: НАДФ-малик-энзимного и НАД-малик-энзимного типа, в то время как ФЕП-карбоксикиназа у ряда видов вовлечена во вторичные пути метаболизма углеводов . При этом анатомо-биохимические особенности таких растений также связаны с экологическими особенностями их роста и развития. Так, например, исследование позволило выявить пять разновидностей Кранц-анатомии листа у С4-представителей семейства Chenopodiaceae.
Исследование биохимических характеристик двудольных С4-растений показало, что в целом описанные показатели были сходными с теми, которые характерны для других С4-видов . В качестве особенностей следует еще раз отметить, что ФЕП-карбоксикиназа не вносит существенного вклада в обмен дикарбоновых кислот в отличие от однодольных трав. С другой стороны, остается во многом неясным присутствие сразу двух декарбоксилирующих ферментов у одного вида (НАД-малик-энзима + ФЕП-карбоксикиназы или НАДФ-малик-энзима + ФЕП-карбоксикиназы). Отчасти это можно объяснить, например, необходимостью синтеза ФЕП (для шикиматного пути), экономией АТФ для обеспечения других процессов и т.п. .
В итоге, было установлено, что среди растений С4-типа преобладают виды с атрипликоидной (характерной для представителей рода Atriplex) анатомией листа и НАДФ-малик-энзимной биохимией.
Особенности «промежуточных» С3-С4-растений
Исследование разных представителей растительного царства дало возможность говорить о специфической фотосинтетической группе, которую стали обозначать, как С3-С4-растения . Характерными чертами этой группы (С3-С4-интермедиатов) являются уникальные особенности газообмена листьев, которые показывают низкую СО2 компенсационную точку и ее криволинейную зависимость от концентрации кислорода в среде. Это обусловлено особенной компартментализацией (местом протекания) фотодыхания в клетках без изменения фотосинтетического метаболизма углерода. Большинство
представителей интермедиатов обладают фотосинтезом С3-типа, а их основной характеристикой является наличие пониженного фотодыхания
Анатомическое строение листьев С3-С4-интермедиатов отличает их от типичных С3- и С4-растений. В отличие от С4-растений у интермедиатов клетки мезофилла не концентрируются вокруг клеток обкладки сосудистых пучков и расположены более рыхло в структуре листа. При этом в клетках интермедиатов содержится большое количество органелл, причем многие из них располагаются в клетках обкладки центрипетально (к центру). Число митохондрий и пероксисом здесь - в четыре раза больше, чем в клетках мезофилла, а митохондрии - вдвое крупнее . В то же время количество плазмодесм между клетками обкладки и мезофилла намного больше, чем у С3-растений, и приближается к показателю, характерному для С4-типа. Это обеспечивает быстрый обмен метаболитами между разными клетками листьев растений.
Ключевой фермент, за счет которого происходит выделение СО2 -глицин-декарбоксилаза, в основном, локализован в клетках обкладки. Особенности взаимного расположения различных органелл в клетках позволяют С3-растениям улавливать до 50 % образуемого (в реакциях преобразования глицина) СО2, в то время как у С3-С4-интермедиатов эта величина может достигать 75 % .
В результате указанные особенности дают возможность интермедиатам проявлять лучшие характеристики в эффективности фотосинтетической ассимиляции СО2, особенно при повышенной температуре, в использовании воды и др., что успешно описывается с помощью математической модели .
Изучение особенностей биохимии показало, что интермедиаты не обладают типичным специфическим распределением ключевых ферментов (ФЕП-карбоксилазы и Рубиско) по разным клеткам, как это имеет место у растений с С4-типом фотосинтеза, показывают неэффективность работы имеющихся компонентов ферментативных систем С4-типа, характеризуются низкой активностью ферментов, связанных с обменом С4-дикарбоновых кислот (ФЕП-карбоксилазы, НАДФ-малат-дегидрогеназы, НАДФ-малик-энзима, пируват-ортофосфат-дикиназы). При этом значительные количества меченого углерода из ассимилируемого 14СО2 оказываются в составе глицина, серина и, особенно, фумарата . В то же время преимущества некоторых С3-С4-растений обусловлены только наличием активного обмена глицина между клетками листьев, о чем говорилось выше. Однако активный обмен углерода должен сопровождаться и активным балансом азота, но картина этого процесса остается неизвестной.
С4-фотосинтез без Кранц-анатомии листа
С4-фотосинтез, обусловленный наличием Кранц-анатомии листа, предполагает, что клеточная стенка между клетками обкладки и мезофилла препятствует потерям СО2, выделяемого при декарбоксилировании С4-метаболита . Однако изучение растительного многообразия, приуроченного к существованию в различных экологических нишах, позволило обнаружить в Центральной Азии растения, в листьях которых многие черты С4-фотосинтеза реализуются в пределах одной клетки без кардинального изменения анатомии листа. При этом известные данные о механизмах концентрирования СО2 у водных растений, а также наличие у некоторых из них С4-фотосинтетического цикла в пределах индивидуальной клетки во многом определяются условиями существования в водной среде.
В отличие от них у наземных растений должна существовать физическая граница (толстая клеточная стенка, мембрана, оболочка), разделяющая клетки, где происходит фотосинтетическая ассимиляция углекислого газа, и клетки, где функционирует цикл Кальвина (рис.).
Упрощенная схема компартментализации С^-фотосинтеза (показаны два типа клеток, между которыми пространственно разделены ассимиляция неорганического углерода и его включение
в продукты цикла Кальвина)
Наземные растения семейства СНвпороШасвав, представленные более чем 1300 видами, часто являются устойчивыми к засухе и засолению, что во многом обеспечивается наличием у них С4-типа фотосинтеза. Среди представителей этого семейства были обнаружены виды, у которых С4-фотосинтез реализуется в пределах одной клетки без
наличия классической Кранц-анатомии листа . Это - Bienertia cycloptera, B. sinuspersici, Suaeda aralocaspica. Они растут в условиях засоления и/или нехватки воды. При этом размеры растений существенно различаются, а именно: если высота B. cycloptera и Suaeda aralocaspica составляет 20 ... 50 см, то растения B. sinuspersici могут достигать 130 ... 160 см.
У этих видов обнаружены два типа топографического распределения органелл внутри клеток. Так, в клетках X aralocaspica различают дистальную и проксимальную части. В дистальной части локализованы агранальные хлоропласты, в которых не синтезируется крахмал, но есть фермент - пируват-ортофосфат-дикиназа для синтеза ФЕП, и здесь (в этой части клетки) осуществляется фиксация СО2 с образованием С4-продукта. В проксимальной части клетки сосредоточены хлоропласты с ферментами цикла Кальвина и малик-энзимом, за счет которого малат преобразуется в пируват с выделением СО2 для его включения в продукты реакций цикла Кальвина с участием Рубиско и других ферментов.
В клетках видов рода Bienertia выделяют центральный компартмент и периферический. В периферической части расположены хлоропласты с малым количеством гран, почти нет митохондрий, но присутствуют ключевые ферменты С4-пути - пируват-ортофосфат-дикиназа и ФЕП-карбоксилаза, обеспечивающие синтез ФЕП и фиксацию углекислого газа, соответственно. В центральной части клетки сосредоточены митохондрии и хлоропласты с развитой гранальной структурой, большим количеством Рубиско и другими ферментами цикла Кальвина . При этом центральный компартмент для избегания потери СО2 окружен вакуолью (вакуолями) .
В то же время у описанных видов наблюдается разное соотношение активностей ключевых ферментов, важных для реализации С3- и С4-путей, что отличает их от настоящих С4-видов .
Еще одним интересным открытием можно признать обнаружение С4-подобного типа фотосинтеза в стеблях и черешках табака , а также в средних жилках листьев арабидопсиса (Arabidopsis ЛаНапа Ь.) -представителей двудольных. Эти особенные черты касаются наличия высокой активности всех ключевых ферментов С4-синдрома, в т.ч. одновременного присутствия высокой активности всех декарбоксилирующих ферментов (НАД- и НАДФ-малик-энзимов, а также ФЕП-карбоксикиназы).
Позднее аналогичное открытие было сделано для представителей однодольных. Так, в клетках, расположенных вокруг центральной жилки проводящего пучка листа типичного С3-растения - риса, была обнаружена подобная повышенная активность указанных выше ферментов . Использование авторами иммуноблот-анализа придает полученным
результатам весомое значение в качестве доказательной базы. Показано, что активность Рубиско в клетках этой части вдвое ниже, чем в других частях листовой пластинки. Активность ФЕП-карбоксилазы увеличена примерно на 30 %. В то же время активность каждого из декарбоксилирующих ферментов (НАД- и НАДФ-малик-энзимов, а также ФЕП-карбоксикиназы) в области средней жилки листа была в шесть и более раз выше. Активность еще одного важнейшего фермента - пируват-ортофосфат-дикиназы - в 7,5 раз превосходила этот показатель для клеток листовой пластинки.
С одной стороны, в ходе обсуждения исследователи интерпретируют такие данные, как один из природных (естественных) путей возможной эволюции и появления С4-фотосинтеза, с другой - такую картину авторы рассматривают как возможность для манипуляций человека по «улучшению» фотосинтеза С3-растений (о чем идут дискуссии с середины 70-х годов прошлого столетия ). Это касается, в первую очередь, такой значимой в мировом масштабе сельскохозяйственной культуры, какой является рис.
При этом авторы исследования значительную долю работы посвятили изучению особенностей работы пигментной системы хлоропластов, которая осуществляет энергетическое обеспечение синтеза органических соединений в ходе фотосинтеза .
С4-фотосинтез в свете эволюции
Появление растений, обладающих С4-типом фотосинтеза, в ходе эволюции, как полагают многие авторы, происходило несколько десятков раз независимыми путями . Основная масса таких растений на сегодняшний день представлена травами (примерно 4600 видов) и осоками (1600 видов), в то время как у двудольных - растения С4-типа представлены примерно 1600 видами, что позволяет говорить о полифилетическом эволюционном происхождении С4-синдрома . При этом эволюционное появление нового пути фотосинтетической ассимиляции СО2 должно было включать ряд этапов, затрагивающих развитие Кранц-анатомии листа, создание СО2-помпы для концентрирования углекислого газа, локализацию и становление С4-цикла, изменения в экспрессии генов с дальнейшей оптимизацией свойств ферментов .
Достаточно ясная и простая теоретическая картина появления С4-фотосинтеза, безусловно, затрагивает и вопросы, касающиеся изменений путей метаболизма других химических элементов и, в первую очередь, азота и серы. Поэтому появление злаковых растений с С4-синдромом могло произойти достаточно давно - примерно 30 млн лет назад, в то время как появление двудольных с таким типом фотосинтеза оценивают в 20 млн лет .
Возникновение С4-фотосинтетического пути рассматривали ранее через появление С3-С4-интермедитов как переходных форм. Изучение многообразия особенностей расположения органелл в клетках мезофилла и обкладки сосудистых пучков, а также ряда ферментов показало, что ключевой механизм в появлении С4-синдрома состоит, по-видимому, в перераспределении между разными клетками глицин-декарбоксилазы, активность которой была резко снижена или потеряна в клетках мезофилла. Косвенным доказательством такого пути является то, что активность этого фермента у С4-растений присутствует только в клетках обкладки. В то же время остается не ясным, почему у таких С3-С4-видов при наличии глицинового «шаттла» (переноса) из одного типа клеток в другой с последующим декарбоксилированием предсказываемые теорией преимущества в скорости ассимиляции СО2, эффективности использования воды и азота не проявляются в условиях обычных концентраций углекислого газа .
Рассмотрение возможностей анатомических изменений структуры листа показывает, что каждая особенность, характерная для С4-синдрома, у С3-растений может меняться независимо (размеры клеток мезофилла и обкладки, толщина листа, расстояние между жилками проводящих пучков), в то время как для С4-фотосинтеза важна координация подобных изменений. С этой позиции переход к С4-растениям многие исследователи рассматривают через возникновение так называемого С2-фотосинтеза, акцентируя внимание на транспорте глицина и его декарбоксилировании в клетках обкладки .
Таким образом, полифилетическое происхождение С4-синдрома стало возможным только благодаря появлению многочисленных характерных черт, что было скоординировано в ходе естественного отбора в меняющихся условиях среды.
Перспективы создания механизмов концентрирования
СО2 у С3-растений
Теоретические изыскания и ряд экспериментальных данных позволили говорить о потенциальной генетической трансформации С3-растений, благодаря которой можно было бы улучшить их (приблизить по фотосинтетическим показателям, устойчивости к факторам среды и продуктивности к С4-растениям) . Позже была экспериментально обоснована возможность работы подобного механизма только при участии оксалоацетат-декарбоксилазы .
В настоящее время решение проблемы повышения эффективности фотосинтеза (и как конечный результат - увеличение продуктивности) С3-растений, представляющих основную массу возделываемых сельскохозяйственных культур, рассматривается в виде нескольких путей.
При этом многие из предложенных решений вызывают обоснованные возражения .
Отдельный интерес в этом аспекте представляет недавняя работа , которая посвящена не только подробному разбору особенностей проявления С4-синдрома для понимания эволюции путей фотосинтеза, но и обоснованию генетического детерминизма эволюции С3-растений в С4-тип. При этом отмечается, что до сих пор генетические механизмы, которые обеспечили появление у растений специфической анатомии листа, характерной для С4-фотосинтеза, остаются неизвестными, и это - главная проблема в попытках биоинженерии по приданию новых характеристик или трансформации большинства важнейших сельскохозяйственных С3-растений в С4-тип.
Особенности генетических механизмов могут состоять в том, что многие черты С4-фотосинтеза реализуются, по-видимому, с помощью многих генов, проявление активности которых имеет малые эффекты, которые только суммарно дают необходимый эффект. При этом пути генетического детерминизма, как показывают данные о различных С3-С4- и С4-растениях, могут быть многообразными, поскольку у представителей отдельных семейств и родов переходные формы Сз^С3-С4^С4 имеют особенные черты и свойства. В этой связи вначале необходимо отследить такие особенности переходов в семействах и родах растений, которые являются таксономически близкими к соответствующим представителям важных сельскохозяйственных культур.
Имеющие место проблемы по трансформации С3-растений в С4-тип описаны в работе Gowik и Westhoff . Авторы отмечают не только необходимость разделения по разным клеткам реакций с участием ФЕП-карбоксилазы и Рубиско, но и затрагивают вопросы о необходимости изменений в экспрессии разных генов. Это связано, например, с тем, что количество Рубиско у С4-растений в несколько раз ниже, чем у С3-растений. При этом механизмы регуляции в клетках каждого типа (мезофилла и обкладки) существенно различаются. Так, у С4-растения Flaveria trinervia в клетках мезофилла присутствует цис-регуляторный элемент, который обозначают как MESOPHYLL EXPRESSION MODULE1. Такой же генетический элемент есть и в генах растений С3-типа этого рода Flaveria, но он не обладает необходимой специфичностью проявления. Небольшая модификация такого элемента достаточна для специфической экспрессии указанного гена .
В противоположность изложенному выше специфическая (для клеток обкладки сосудистых пучков) экспрессия одного из генов, кодирующих малую субъединицу Рубиско, FbRbcSl, регулируется в основном на посттранскрипционном уровне. При этом кажется, что стабильность транскриптов этого гена в клетках мезофилла и обкладки различная .
Таким образом, трансформация Сз-растений в С4-тип во многом связана с изменением регуляции активности разных генов. При этом ключевую роль играют, по-видимому, GOLDEN2-LIKE транскрипционные факторы GLK1 и GLK2, которые характерны для всех наземных растений . У растения Arabidopsis они контролируют экспрессию более 100 генов, многие из которых связаны с фотосинтетическим процессом, в то время как у кукурузы (С4-растения) наблюдали специфичную для клеток обкладки экспрессию генов этой группы . Такие результаты дали основание авторам сделать заключение о том, что эти белки являются важными для генной регуляции в образовании клеток мезофилла и обкладки сосудистых пучков. Однако дальнейшие шаги могут касаться оптимизации количества и свойств многих ферментов.
Таким образом, эволюция фотосинтеза представляется весьма сложной и многообразной, причем проблемы биоинженерии и управления продуктивностью растений с помощью генетических манипуляций, по-видимому, имеют перспективы решения с учетом накопленных знаний о путях регуляции морфологических и биохимических характеристик, важных для проявления более эффективного С4-пути фотосинтеза.
Список литературы
1. Хелдт Г.-В. Биохимия растений. М. : БИНОМ. Лаборатория знаний, 2011. 471 с.
2. Tolbert N. E. The C2 oxidative photosynthetic carbon cycle // Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997. V. 48. P. 1-25.
3. Lundgren M.R., Osborne C.P., Christin P.-A. Deconstructing Kranz anatomy to understand C4 evolution // J. Exp. Bot. 2014. V. 65 (13). P. 33573369.
4. Proof of C4 photosynthesis without Kranz anatomy in Bienertia cycloptera (Chenopodiaceae) / E.V. Voznesenskaya // The Plant Journal. 2002. V. 31 (5). P. 649-662.
5. Sage R.F. C4 photosynthesis in terrestrial plants does not require Kranz anatomy // TRENDS in Plant Science. 2002. V. 7 (7). P. 283-285.
6. Wickramage A.T. Single cell C4 photosynthesis. 2008. https://www.slideshare.net/anupathisalwickramage/single-cell-c4photosynthesis.
7. The existence of C-4-bundle-sheath-like photosynthesis in the mid-vein of C-3 rice / W.J. Shen // Rice. 2016. V. 9. No 20. DOI: 10.1186/s12284-016-0094-5.
8. Gowik U., Westholl P. The path from C3 to C4 photosynthesis // Plant Physiol. 2011. V. 155. P. 56-63.
9. Muhaidat R., Sage R.F., Dengler N.G. Diversity of Kranz anatomy and biochemistry in C4 eudicots // Amer. J. Bot. 2007. V.94 (3). P. 362-381.
10. C2 photosynthesis generates about 3-fold elevated leaf CO2 levels in the C3-C4 intermediate species Flaveria pubescens / O. Keerberg //
Journal of Experimental Botany. 2014. V. 65. No. 13. P. 3649-3656. doi:10.1093/jxb/eru239.
11. Glycine decarboxylase is confined to the bundle-sheath cells of leaves of C3-C4 intermediate species / C.M. Hylton // Planta. 1988. V. 175. P. 452-459.
12. Sage R.F., Sage T.L., Kocacinar F. Photorespiration and the evolution of C4 photosynthesis // Annual Review of Plant Biology. 2012. V. 63. P. 19-47.
13. Phenotypic landscape inference reveals multiple evolutionary paths to C4 photosynthesis / B.P. Williams // eLife 2. 2013. e00961. doi.org/10.7554/eLife.00961.
14. Эдвардс Дж., Уокер Д. Фотосинтез С3- и С4-растений: механизмы и регуляция. М.: Мир, 1986. 590 с.
15. Monson R.K. CO2 assimilation in C3-C4-intermediate plants // Photosynthesis: Physiology and Metabolism / R.C. Leegood, T.D. Sharkey, S. von Caemmerer (eds). Kluwer Acadimic Publishers. 2000. P. 533-550.
16. Sage R.F. The evolution of C4 photosynthesis // New Phytologist. 2004. V. 161. P. 341-370.
17. Hunt S., Smith A.M., Woolhouse H.W. Evidence for a light-dependent system for reassimilation of photorespiratory which does not include a cycle, in the intermediate species Moricandia arvensis // Planta. 1987. V. 171. P. 227- 234.
18. von Caemmerer S. A model of photosynthetic assimilation and carbon-isotope discrimination in leaves of certain intermediates // Planta. 1989. V. 178. P. 463-474.
19. Иванищев В.В. Об определении эффективности карбоксилирования у растений, обладающих механизмом концентрирования СО2 // Физиология растений. 1992. Т. 39. № 3. С. 437444.
20. Hibberd J.M., Quick W.P. Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants // Nature. 2002. V. 415 (6870). P. 451-454. doi:10.1038/415451a.
21. C4 acid decarboxylases required for C4 photosynthesis are active in the mid-vein of the C4 species Arabidopsis thaliana, and are important in sugar and amino acid metabolism / N.J. Brown // Plant J. 2010. V. 61(1). P. 122-133. doi:10.1111/j.1365-313X.2009.04040.x.
22. Магомедов И.М. К вопросу об истории открытия С4-фотосинтеза. Современное состояние проблемы // Успехи современного естествознания. 2015. № 1 (часть 6). С. 962-965.
23. Насыров Ю.С. Генетика фотосинтеза и селекция. М.: Знание, 1982. 64 с.
24. Абдуллаев А., Горенкова Л.Г., Иванищев В.В. О распределении некоторых ферментов метаболизма С4-кислот в хлоропластах ржи // Физиология растений. 1989. Т. 36. № 4. С. 665-668.
25. Иванищев В.В. Биологическое значение метаболизма оксало-ацетата в хлоропластах С3- растений // Физиология растений. 1997. Т. 44. № 3. С. 462-470.
26. Meyer V., Griffiths H. Origins and diversity of eucariotic CO2-concentrating mechanisms: lessons for the future // J. Exp. Bot. 2013. V. 64. No 3. P. 769-786.
27. Иванищев В.В. Проблемы фотосинтетической ассимиляции неорганического углерода высшими растениями // Вестник ГОУ ДПО ТО "ИПК И ППРО ТО". Тульское образовательное пространство. 2017. № 3 (в печати).
28. Evolution and function of a cis-regulatory module for mesophyllspecific gene expression in the C4 dicot Flaveria trinervia / M. Akyildiz // Plant Cell. 2007. V. 19. P. 3391-3402.
29. Patel M., Siegel A.J., Berry J.O. Untranslated regions of FbRbcS1 mRNA mediate bundle sheath cell-specific gene expression in leaves of a C4 plant // J. Biol Chem. 2006. V. 281. P. 25485-25491.
30. Waters M.T., Langdale J.A. The making of a chloroplast // EMBO J. 2009. V. 28. P. 2861-2873.
Иванищев Виктор Васильевич, д-р биол. наук, ст. науч. сотрудник, зав. кафедрой, [email protected], Россия, Тула, Тульский государственный педагогический университет им. Л.Н. Толстого
EVOLUTIONARY ASPECTS OF C4-PHOTOSYNTHESIS V.V. Ivanishchev
The review deals with the variety of manifestations of photosynthetic assimilation of carbon dioxide in plants and the evolutionary aspects of the C4-type photosynthesis. It is shown that over the last decade and a half, experimental data have been obtained that significantly change our understanding of the ways of the onset of C4 photosynthesis and the realization of its specific properties in a number of typical C3 plants. The prospects of application such knowledge for developing a strategy for improving the efficiency of photosynthesis of C3 plants are discussed in order to increase their productivity.
Key words: plants, assimilation of inorganic carbon, C3-C4-photosynthesis, C4-photosynthesis, evolutionary aspects, plant productivity.
Ivanishchev Viktor Vasiljevich, Doctor of Biology, Senior Researcher, head of chair, [email protected], Russia, Tula, Tula State Lev Tolstoy Pedagogical University
Поскольку в цикле Кальвина первичными продуктами включения неорганического углерода в органический являются трехуглеродные соединения, данный процесс носит название С-3 путь фотосинтеза.
Важно заметить, что для синтеза одной молекулы глюкозы должно произойти шесть оборотов цикла Кальвина. В каждом обороте используются три молекулы АТФ (две для активирования двух молекул фосфоглицериновой кислоты и одна при регенерации рибулезодифосфата) и две молекулы НАДФ. Н 2 для восстановления кислоты в альдегид. Таким образом для синтеза одной молекулы глюкозы крайне важно потратить 12 молекул НАДФ. Н 2 и 18 молекул АТФ.
Важно отметить, что физиологическое значение цикла Кальвина состоит не только в акцепции углекислого газа, но и в создании массы углеводных соединений, которые идут как на синтез запасных веществ, так и на создание компонентов хлоропласта и текущий метаболизм клетки. .
Большинство растений усваивает неорганический углерод именно по пути цикла Кальвина. При этом довольно большая группа растений (около 500 видов) тропического происхождения выработала в процессе эволюции некоторую модификацию процесса, усваивая неорганический углерод путем образования в результате его акцепции четырехуглеродных соединений. Это растения, приспособившиеся к фотосинтезу в условиях повышенной температуры воздуха и избыточной освещенности, а также пониженной влажности почвы (засухи). Из культурных растений обладают таким метаболитическим процессом кукуруза, просо, сорго, сахарный тростник. У ряда сорных растений также наблюдается именно эта особенность метаболизма (свинорой, просо куриное, щирица) и т.д.
Особенностью анатомического строения таких растений является наличие фотосинтезирующих клеток двух типов, которые располагаются в виде концентрических кругов - радиально расположенные вокруг проводящих пучков клетки обкладочной паренхимы и мезофилла. Этот тип анатомического строения принято называть кранц-типом (от немецкого Кranz -венок).
Этот тип метаболизма был изучен в 60-е годы прошлого века, большую роль сыграли при этом исследования советских ученых Карпилова, Незговоровой, Тарчевского, а также австралийских ученых Хэтча и Слэка. Именно они предложили законченную схему цикла, в связи с этим принято данный процесс называть также циклом Хэтча-Слэка-Карпилова.
Процесс происходит в два этапа: поступающий в мезофилл СО 2 вступает в соединение с трехуглеродным соединением (ФЕП) - фосфоенолпировиноградной кислотой - которая превращается в четырехуглеродное соединение. Это и есть ключевой момент процесса, из-за которого он и получил свое название, так как неорганический углерод, акцептируясь трехуглеродным соединением, превращается в четырехуглеродное соединение. Учитывая зависимость оттого в какое именно четырехуглеродное соединение превращается неорганический углерод различают три группы растений:
НАДФ-МДГ образуют яблочную кислоту при участии фермента малатдегидрогеназы, а затем пировиноградной кислоты,
НАД-МДГ образуют аспарагиновую кислоту и аланин,
ФЕП-КК образуют аспарагиновую кислоту и фосфоенолпировиноградную кислоту.
Наиболее значимые для сельского хозяйства растения относятся к НАДФ-МДГ типу.
После образования четырехуглеродного соединения происходит его перемещение во внутренние клетки обкладочной паренхимы и расщепление или декарбоксилирование этой молекулы. Отделившаяся карбоксильная группа в виде CОО - входит в цикл Кальвина, а оставшаяся трехуглеродная молекула - ФЕП - возвращается опять в клетки мезофилла.
Такой путь фиксации углекислого газа позволяет растениям накапливать в виде органических кислот запас углерода, проводить процесс фотосинтеза в наиболее жаркое время дня при сокращении потерь воды на транспирацию за счёт закрытия устьиц. Эффективность использования воды такими растениями в два раза больше, чем у растений, происходящих из умеренных широт.
Для С 4 -растений характерны отсутствие обратного потока углекислого газа при фотодыхании и повышенный уровень синтеза и накопления органических веществ.