Адаптированные растения. Основные этапы эволюции растений и животных. Типы растений по отношению к водному режиму
Теперь, когда мы познакомились с отличительными признаками четырех основных групп растений, а именно моховидных, папоротниковидных, голосеменных и покрытосеменных (цветковых), нам легче представить эволюционный прогресс, сделанный растениями в процессе адаптации к жизни на суше.
Проблемы
Пожалуй, наитруднейшей проблемой, которую надо было как-то преодолеть, чтобы перейти от водного образа жизни к наземному, была проблема обезвоживания . Любое растение, незащищенное тем или иным способом, например не покрытое восковой кутикулой, очень скоро высохнет и несомненно погибнет. Даже если преодолеть эту трудность, останутся другие нерешенные проблемы. И прежде всего вопрос о том, как успешно осуществить половое размножение. У первых растений в размножении участвовали мужские гаметы, способные приблизиться к женским гаметам, только плавая в воде.
Обычно считают, что первые растения, освоившие сушу, произошли от зеленых водорослей, у отдельных из наиболее эволюционно продвинутых представителей которых появились репродуктивные органы, а именно архегонии (женские) и антеридии (мужские); в этих органах были спрятаны, а, следовательно и защищены гаметы. Это обстоятельство и ряд других вполне определенных приспособлений, помогающих избежать высыхания, позволили некоторым представителям зеленых водорослей завладеть сушей.
Одна из важнейших эволюционных тенденций у растений – это постепенно увеличивающаяся независимость их от воды.
Ниже перечислены те основные трудности, которые связаны с переходом от водного к наземному существованию.
- Обезвоживание. Воздух – это среда, способствующая высыханию, а вода необходима для жизни по целому ряду причин (разд. 3.1.2). Следовательно, возникает необходимость в приспособлениях для получения и запасания воды.
- Размножение. Нежные половые клетки должны быть защищены, а подвижные мужские гаметы (спермии) могут встретиться с женскими гаметами только в воде.
- Опора. В отличие от воды воздух не может служить опорой растениям.
- Питание. Растениям необходимы свет и диоксид углерода (CO 2) для фотосинтеза, поэтому хотя бы часть растения должна возвышаться над землей. Однако минеральные соли и вода находятся в почве или на ее поверхности, и, чтобы эффективно использовать эти вещества, часть растения должна находиться в земле и расти в темноте.
- Газообмен. Для фотосинтеза и дыхания нужно, чтобы обмен диоксида углерода и кислорода происходил не с окружающим раствором, а с атмосферой.
- Факторы окружающей среды. Вода, особенно, когда ее так много, как, скажем, в озере или в океане, обеспечивает высокое постоянство условий окружающей среды. Наземная же среда обитания в гораздо большей степени характеризуется изменчивостью таких важных факторов, как температура, интенсивность освещения, концентрация ионов и pH.
Печеночники и мхи
Мхи хорошо приспособились к распространению спор в наземных условиях: оно зависит от высыхания коробочки и рассеивания мелких легких спор ветром. Однако эти растения все еще находятся в зависимости от воды по следующим причинам.
- Вода необходима им для размножения, поскольку спермии должны подплывать к архегониям. У этих растений возникли адаптации, позволяющие им высвобождать спермии только во влажной среде, потому что только в такой среде вскрываются антеридии. Эти растения частично приспособились к наземной жизни, поскольку гаметы у них образуются в защитных структурах – антеридиях и архегониях.
- У них нет специальных опорных тканей, и поэтому рост растения вверх ограничен.
- У моховидных нет корней, способных далеко проникать в субстрат, и они могут жить только там, где на поверхности почвы или в ее верхних слоях имеется достаточно влаги и минеральных солей. Однако у них имеются ризоиды, которыми они прикрепляются к грунту; это – одна из адаптаций к жизни на твердом субстрате.
2.4. Печеночники и мхи часто называют амфибиями (земноводными) растительного мира. Объясните вкратце, почему.
Папоротники
2.5. Папоротники лучше адаптировались к жизни на суше, чем печеночники и мхи. В чем это проявляется?
2.6. По каким важным признакам мхи, папоротники и печеночники плохо адаптировались к жизни на суше?
Семенные растения – хвойные и цветковые
Одна из основных трудностей, с которой сталкиваются растения на суше, связана с уязвимостью гаметофитного поколения. Например, у папоротников гаметофит – это нежный заросток, который образует мужские гаметы (спермии), нуждающиеся в воде, чтобы достичь яйцеклетки. Однако у семенных растений гаметофит защищен и сильно редуцирован.
Семенные растения обладают тремя важными преимуществами: во-первых, разноспоровостью; во-вторых, появлением неплавающих мужских гамет и, в-третьих, образованием семян.
РАЗНОСПОРОВОСТЬ И НЕПЛАВАЮЩИЕ МУЖСКИЕ ГАМЕТЫ.
Рис. 2.34. Обобщенная схема жизненного цикла растений, отражающая чередование поколений. Обратите внимание на наличие гаплоидных (n) и диплоидных (2n) стадий. Гаметофит всегда гаплоидный и всегда образует гаметы путем митотического деления. Спорофит всегда диплоидный и всегда образует споры в результате мейотического деления.
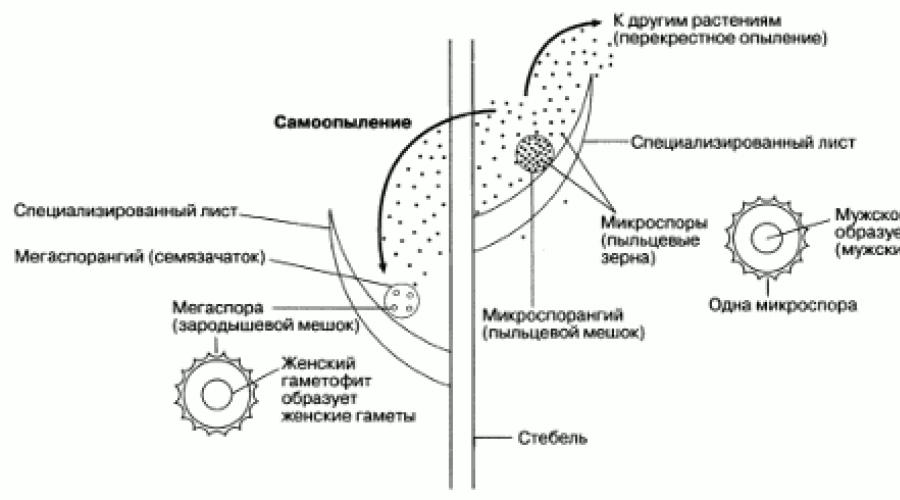
Очень важную роль в эволюции растений сыграло возникновение некоторых папоротников и их близких родичей, образующих споры двух типов. Явление это называют разноспоровостью , а растения – разноспоровыми. Все семенные растения относятся к разноспоровым. Они образуют крупные споры, называемые мегаспорами , в спорангиях одного типа (мегаспорангиях) и мелкие споры, называемые микроспорами, – в спорангиях другого типа (микроспорангиях). Прорастая, споры образуют гаметофиты (рис. 2.34). Мегаспоры развиваются в женские гаметофиты, микроспоры – в мужские. У семенных растений гаметофиты, образуемые мегаспорами и микроспорами, очень малы по размерам и никогда не высвобождаются из спор. Таким образом, гаметофиты оказываются защищенными от высыхания, что представляет собой важное эволюционное достижение. Однако спермии из мужского гаметофита все еще должны перемещаться к женскому гаметофиту, что значительно облегчается рассеиванием микроспор. Будучи очень мелкими, они могут образовываться в больших количествах и разноситься ветром далеко от родительского спорофита. Случайно они могут оказаться в тесной близости от мегаспоры, которая у семенных растений не отделяется от родительского спорофита (рис. 2.45). Именно таким путем и происходит опыление у растений, пыльцевые зерна которых представляют собой микроспоры. В пыльцевых зернах образуются мужские гаметы.

Рис. 2.45. Схематическое изображение основных элементов разноспоровости и опыления.
У семенных растений возникло еще одно эволюционное преимущество. Мужским гаметам не нужно больше подплывать к женским гаметам, поскольку у семенных растений появились пыльцевые трубки. Они развиваются из пыльцевых зерен и растут в направлении женских гамет. По этой трубке мужские гаметы достигают женской гаметы и оплодотворяют ее. Плавающие спермии больше не образуются, в оплодотворении участвуют только мужские ядра.
Следовательно, у растений выработался механизм оплодотворения, независимый от воды. Это и послужило одной из причин, по которой семенные растения столь превзошли другие растения в освоении суши. Первоначально опыление происходило только с помощью ветра – процесс довольно случайный, сопровождающийся большими потерями пыльцы. Однако уже на ранних этапах эволюции примерно 300 млн. лет назад в каменноугольном периоде, появились летающие насекомые, а с ними и возможность более эффективного опыления. Цветковые растения широко используют опыление насекомыми, тогда как у хвойных все еще преобладает опыление ветром.
СЕМЕНА. У ранних разноспоровых растений мегаспоры высвобождались из родительского спорофита подобно микроспорам. У семенных же растений мегаспоры не отделяются от родительского растения, оставаясь в мегаспорангиях, или семязачатках (рис. 2.45). Семязачаток содержит женскую гамету. После оплодотворения женской гаметы семязачаток называют уже семенем . Таким образом, семя – это оплодотворенный семязачаток. Наличие семязачатка и семени дает определенные преимущества семенным растениям.
- Женский гаметофит защищен семязачатком. Он полностью зависит от родительского спорофита и в отличие от свободно живущего гаметофита нечувствителен к обезвоживанию.
- После оплодотворения в семени образуется запас питательных веществ, получаемых гаметофитом от родительского спорофитного растения, от которого он по-прежнему не отделен. Этот запас используется развивающейся зиготой (следующим спорофитным поколением) после прорастания семени.
- Семена предназначены для того, чтобы переживать неблагоприятные условия, и остаются в состоянии покоя до тех пор, пока условия не станут благоприятными для прорастания.
- У семян могут развиваться различные приспособления, облегчающие их распространение.
Семя представляет собой сложную структуру, в которой собраны клетки трех поколений – родительского спорофита, женского гаметофита и зародыша следующего спорофитного поколения. Родительский спорофит дает семени все, что нужно для жизни, и только после того, как семя полностью созреет, т.е. накопит запас питательных веществ для зародыша спорофита, оно отделяется от родительского спорофита.
2.7. Шансы для выживания и развития пыльцевых зерен (микроспор), переносимых ветром, намного меньше, чем для спор Dryopteris. Почему?
2.8. Объясните, почему мегаспоры крупные, а микроспоры мелкие.
2.7.7. Краткое перечисление адаптаций семенных растений к жизни на суше
Основные преимущества семенных растений над всеми остальными сводятся к следующему.
- Гаметофитное поколение сильно редуцировано и полностью зависит от хорошо приспособленного к жизни на суше спорофита, внутри которого гаметофит всегда защищен. У других растений гаметофит очень легко высыхает.
- Оплодотворение происходит независимо от воды. Мужские гаметы неподвижны и разносятся внутри пыльцевых зерен ветром или насекомыми. Окончательный перенос мужских гамет к женским происходит с помощью пыльцевой трубки.
- Оплодотворенные семязачатки (семена) остаются некоторое время на родительском спорофите, от которого они получают защиту и пищу прежде, чем будут развеяны.
- У многих семенных растений наблюдается вторичный рост с отложением больших количеств древесины, несущей опорную функцию. Такие растения вырастают в деревья и кустарники, способные эффективно конкурировать в борьбе за свет и другие ресурсы.
Некоторые из важнейших эволюционных тенденций приводятся в обобщенном виде на рис. 2.33. У семенных растений имеются и другие признаки, присущие растениям не только этой группы, но также выполняющие роль адаптаций к жизни на суше.

Рис. 2.33. Систематика растений и некоторые основные тенденции в эволюции растений.
- Настоящие корни обеспечивают извлечение влаги из почвы.
- Растения защищены от высыхания эпидермисом с водонепроницаемой кутикулой (или пробкой, образующейся после вторичного роста).
- Эпидермис наземных частей растения, особенно листьев, пронизан множеством мельчайших щелей, называемых устьицами , через которые осуществляется газообмен между растением и атмосферой.
- У растений имеются и специализированные адаптации к жизни в жарких засушливых условиях (гл. 19 и 20).
Приспособленность онтогенеза растений к условиям среды является результатом их эволюционного развития (изменчивости, наследственности, отбора). На протяжении филогенеза каждого вида растений в процессе эволюции выработались определенные потребности индивидуума к условиям существования и приспособленность к занимаемой им экологической нише. Влаголюбие и теневыносливость, жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в ходе эволюции в результате длительного действия соответствующих условий. Так, теплолюбивые растения и растения короткого дня характерны для южных широт, менее требовательные к теплу и растения длинного дня - для северных.
В природе в одном географическом регионе каждый вид растений занимает экологическую нишу, соответствующую его биологическим особенностям: влаголюбивые - ближе к водоемам, теневыносливые - под пологом леса и т. д. Наследственность растений формируется под влиянием определенных условий внешней среды. Важное значение имеют и внешние условия онтогенеза растений.
В большинстве случаев растения и посевы (посадки) сельскохозяйственных культур, испытывая действие тех или иных неблагоприятных факторов, проявляют устойчивость к ним как результат приспособления к условиям существования, сложившимся исторически, что отмечал еще К. А. Тимирязев.
1. Основные среды жизни.
При изучении окружающей среды (среды обитания растений и животных и производственной деятельности человека) выделяют следующие ее основные составляющие: воздушную среду; водную среду (гидросферу); животный мир(человек, домашние и дикие животные, в том числе рыбы и птицы); растительный мир (культурные и дикие растения в том числе растущие в воде);почву(растительный слой);недра(верхняя часть земной коры, в пределах которой возможна добыча полезных ископаемых); климатическую и акустическую среду.
Воздушная среда может быть наружной, в которой большинство людей проводят меньшую часть времени (до 10-15%), внутренней производственной (в ней человек проводит до 25-30% своего времени) и внутренней жилой, где люди пребывают большую часть времени (до 60-70% и более).
Наружный воздух у поверхности земли содержит по объему: 78,08% азота; 20,95% кислорода; 0,94% инертных газов и 0,03% углекислого газа. На высоте 5 км содержание кислорода остается тем же, а азота увеличивается до 78,89%. Часто воздух у поверхности земли имеет различные примеси, особенно в городах: там он содержит более 40 ингредиентов, чуждых природной воздушной среде. Внутренний воздух в жилищах, как правило, имеет
повышенное содержание углекислого газа, а внутренний воздух производственных помещений обычно содержит примеси, характер которых определяется технологией производства. Среди газов выделяется водяной пар, который попадает в атмосферу в результате испарений с Земли. Большая его часть (90%) сосредоточена в самом нижнем пятикилометровом слое атмосферы, с высотой его количество очень быстро уменьшается. Атмосфера содержит много пыли, которая попадает туда с поверхности Земли и частично из космоса. При сильных волнениях ветры подхватывают водяные брызги из морей и океанов. Так попадают в атмосферу из воды частицы соли. В результате извержения вулканов, лесных пожаров, работы промышленных объектов и т.д. воздух загрязняется продуктами неполного сгорания. Больше всего пыли и других примесей в приземном слое воздуха. Даже после дождя в 1 см содержится около 30 тыс. пылинок, а в сухую погоду их в сухую погоду их в несколько раз больше.
Все эти мельчайшие примеси влияют на цвет неба. Молекулы газов рассеивают коротковолновую часть спектра солнечного луча, т.е. фиолетовые и синие лучи. Поэтому днем небо голубого цвета. А частицы примесей, которые значительно крупней молекул газов, рассеивают световые лучи почти всех длин волн. Поэтому, когда воздух запылен или в нем содержатся капельки воды, небо становится белесоватым. На больших высотах небо темно-фиолетовое и даже черное.
В результате происходящего на Земле фотосинтеза растительность ежегодно образует 100 млрд. т. органических веществ (около половины приходится на долю морей и океанов), усваивая при этом около 200 млрд. т. углекислого газа и выделяя во внешнюю среду около 145 млрд.т. свободного кислорода, полагают, что благодаря фотосинтезу образуется весь кислород атмосферы. О роли в этом круговороте зеленых насаждений говорят следующие данные: 1 га зеленых насаждений в среднем за 1 час очищает воздух от 8 кг углекислого газа (выделяемого за это время при дыхании 200 человек). Взрослое дерево за сутки выделяет 180 литров кислорода, а за пять месяцев (с мая по сентябрь) оно поглощает около 44 кг углекислого газа.
Количество выделяемого кислорода и поглощаемого углекислого газа зависит от возраста зеленых насаждений, видового состава, плотности посадки и других факторов.
Не меньшее значение имеют и морские растения - фитопланктон(в основном водоросли и бактерии), высвобождающие путем фотосинтеза кислород.
Водная среда включает поверхностные и подземные воды. Поверхностные воды в основном сосредоточены в океане, содержанием 1 млрд. 375 млн. кубических километров - около 98% всей воды на Земле. Поверхность океана (акватория) составляет 361 млн. квадратных километров. Она примерно в 2,4 раза больше площади суши--территории, занимающей 149 млн. квадратных километров. Вода в океане соленая, причем большая ее часть (более 1 млрд. кубических километров) сохраняет постоянную соленость около 3,5% и температуру, примерно равную 3,7є С. Заметные различия в солености и температуре наблюдаются почти исключительно в поверхностном слое воды, а также в окраинных и особенно в средиземных морях. Содержание растворенного кислорода в воде существенно уменьшается на глубине 50-60 метров.
Подземные воды бывают солеными, солоноватыми (меньшей солености) и пресными; существующие геотермальные воды имеют повышенную температуру (более 30єС).
Для производственной деятельности человечества и его хозяйственно-бытовых нужд требуется пресная вода, количество которой составляет всего лишь 2,7% общего объема воды на Земле, причем очень малая ее доля (всего 0,36%) имеется в легкодоступных для добычи местах. Большая часть пресной воды содержится в снегах и пресноводных айсбергах, находящихся в районах в основном Южного полярного круга.
Годовой мировой речной сток пресной воды составляет 37,3 тыс. кубических километров. Кроме того, может использоваться часть подземных вод, равная 13 тыс. кубическим километрам. К сожалению, большая часть речного стока в России, составляющая около 5000 кубических километров, приходится на малоплодородные и малозаселенные северные территории.
Климатическая среда является важным фактором, определяющим развитие различных видов животного, растительного мира и его плодородие. Характерной особенностью России является то, что большая часть ее территории имеет значительно более холодный климат, чем в других странах.
Все рассмотренные составляющие окружающей среды входят в
БИОСФЕРУ: оболочку Земли, включающую часть атмосферы, гидросферу и верхнюю часть литосферы, которые взаимно связанны сложными биохимическими циклами миграции вещества и энергии, геологическую оболочку Земли, населенную живыми организмами. Верхний предел жизни биосферы ограничен интенсивной концентрацией ультрафиолетовых лучей; нижний - высокой температурой земных недр (свыше100`С). Крайних пределов ее достигают только низшие организмы - бактерии.
Адаптация (приспособление) растения к конкретным условиям среды обеспечивается за счет физиологических механизмов (физиологическая адаптация), а у популяции организмов (вида) - благодаря механизмам генетической изменчивости, наследственности и отбора (генетическая адаптация). Факторы внешней среды могут изменяться закономерно и случайно. Закономерно изменяющиеся условия среды (смена сезонов года) вырабатывают у растений генетическую приспособленность к этим условиям.
В естественных для вида природных условиях произрастания или возделывания растения в процессе своего роста и развития часто испытывают воздействие неблагоприятных факторов внешней среды, к которым относят температурные колебания, засуху, избыточное увлажнение, засоленность почвы и т. д. Каждое растение обладает способностью к адаптации в меняющихся условиях внешней среды в пределах, обусловленных его генотипом. Чем выше способность растения изменять метаболизм в соответствии с окружающей средой, тем шире норма реакции данного растения и лучше способность к адаптации. Это свойство отличает устойчивые сорта сельскохозяйственных культур. Как правило, несильные и кратковременные изменения факторов внешней среды не приводят к существенным нарушениям физиологических функций растений, что обусловлено их способностью сохранять относительно стабильное состояние при изменяющихся условиях внешней среды, т. е. поддерживать гомеостаз. Однако резкие и длительные воздействия приводят к нарушению многих функций растения, а часто и к его гибели.
При действии неблагоприятных условий снижение физиологических процессов и функций может достигать критических уровней, не обеспечивающих реализацию генетической программы онтогенеза, нарушаются энергетический обмен, системы регуляции, белковый обмен и другие жизненно важные функции растительного организма. При воздействии на растение неблагоприятных факторов (стрессоров) в нем возникает напряженное состояние, отклонение от нормы - стресс. Стресс - общая неспецифическая адаптационная реакция организма на действие любых неблагоприятных факторов. Выделяют три основные группы факторов, вызывающих стресс у растений: физические - недостаточная или избыточная влажность, освещенность, температура, радиоактивное излучение, механические воздействия; химические - соли, газы, ксенобиотики (гербициды, инсектициды, фунгициды, промышленные отходы и др.); биологические - поражение возбудителями болезней или вредителями, конкуренция е другими растениями, влияние животных, цветение, созревание плодов.
Сила стресса зависит от скорости развития неблагоприятной для растения ситуации и уровня стрессирующего фактора. При медленном развитии неблагоприятных условий растение лучше приспосабливается к ним, чем при кратковременном, но сильном действии. В первом случае, как правило, в большей степени проявляются специфические механизмы устойчивости, во втором - неспецифические.
В неблагоприятных природных условиях устойчивость и продуктивность растений определяются рядом признаков, свойств и защитно-приспособительных реакций. Различные виды растений обеспечивают устойчивость и выживание в неблагоприятных условиях тремя основными способами: с помощью механизмов, которые позволяют им избежать неблагоприятных воздействий (состояние покоя, эфемеры и др.); посредством специальных структурных приспособлений; благодаря физиологическим свойствам, позволяющим им преодолеть пагубное влияние окружающей среды.
Однолетние сельскохозяйственные растения в умеренных зонах, завершая свой онтогенез в сравнительно благоприятных условиях, зимуют в виде устойчивых семян (состояние покоя). Многие многолетние растения зимуют в виде подземных запасающих органов (луковиц или корневищ), защищенных от вымерзания слоем почвы и снега. Плодовые деревья и кустарники умеренных зон, защищаясь от зимних холодов, сбрасывают листья.
Защита от неблагоприятных факторов среды у растений обеспечивается структурными приспособлениями, особенностями анатомического строения (кутикула, корка, механические ткани и т. д.), специальными органами защиты (жгучие волоски, колючки), двигательными и физиологическими реакциями, выработкой защитных веществ (смол, фитонцидов, токсинов, защитных белков).
К структурным приспособлениям относятся мелколистность и даже отсутствие листьев, воскообразная кутикула на поверхности листьев, их густое опущение и погруженность устьиц, наличие сочных листьев и стеблей, сохраняющих резервы воды, эректоидность или пониклость листьев и др. Растения располагают различными физиологическими механизмами, позволяющими приспосабливаться к неблагоприятным условиям среды. Это сам-тип фотосинтеза суккулентных растений, сводящий к минимуму потери воды и крайне важный для выживания растений в пустыне и т. д.
2. Приспособление у растений
Холодостойкость растений
Устойчивость растений к низким температурам подразделяют на холодостойкость и морозоустойчивость. Под холодостойкостью понимают способность растений переносить положительные температуры несколько выше Оє С. Холодостойкость свойственна растениям умеренной полосы (ячмень, овес, лен, вика и др.). Тропические и субтропические растения повреждаются и отмирают при температурах от 0є до 10є С (кофе, хлопчатник, огурец и др.). Для большинства же сельскохозяйственных растений низкие положительные температуры негубительны. Связано это с тем, что при охлаждении ферментативный аппарат растений не расстраивается, не снижается устойчивость к грибным заболеваниям и вообще не происходит заметных повреждений растений.
Степень холодостойкости разных растений неодинакова. Многие растения южных широт повреждаются холодом. При температуре 3 °С повреждаются огурец, хлопчатник, фасоль, кукуруза, баклажан. Устойчивость к холоду у сортов различна. Для характеристики холодостойкости растений используют понятие температурный минимум, при котором рост растений прекращается. Для большой группы сельскохозяйственных растений его величина составляет 4 °С. Однако многие растения имеют более высокое значение температурного минимума и соответственно они менее устойчивы к воздействию холода.
Приспособление растений к низким положительным температурам.
Устойчивость к низким температурам - генетически детерминированный признак. Холодостойкость растений определяется способностью растений сохранять нормальную структуру цитоплазмы, изменять обмен веществ в период охлаждения и последующего повышения температуры на достаточно высоком уровне.
Морозоустойчивость растений
Морозоустойчивость - способность растений переносить температуру ниже О °С, низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Морозы в зимний период с температурой ниже -20 °С обычны для значительной части территории России. Воздействию морозов подвергаются однолетние, двулетние и многолетние растения. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних - клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани этих растений могут замерзать, однако растения не погибают.
Замерзание растительных клеток и тканей и происходящие при этом процессы.
Способность растений переносить отрицательные температуры определяется наследственной основой данного вида растений, однако морозоустойчивость одного и того же растения зависит от условий, предшествующих наступлению морозов, влияющих на характер льдообразования. Лед может образовываться как в протопласте клетки, так и в межклеточном пространстве. Не всякое образование льда приводит клетки растения к гибели.
Постепенное снижение температуры со скоростью 0,5-1 °С/ч приводит к образованию кристаллов льда прежде всего в межклеточниках и первоначально не вызывают гибели клеток. Однако последствия этого процесса могут быть губительными для клетки. Образование льда в протопласте клетки, как правило, происходит при быстром понижении температуры. Происходит коагуляция белков протоплазмы, кристаллами образовавшегося в цитозоле льда повреждаются клеточные структуры, клетки погибают. Убитые морозом растения после оттаивания теряют тургор, из их мясистых тканей вытекает вода.
Морозоустойчивые растения обладают приспособлениями, уменьшающими обезвоживание клеток. При понижении температуры у таких растений отмечаются повышение содержания Сахаров и других веществ, защищающих ткани (криопротекторы), это прежде всего гидрофильные белки, моно- и олигосахариды; снижение оводненности клеток; увеличение количества полярных липидов и снижение насыщенности их жирнокислотных остатков; увеличение количества защитных белков.
На степень морозоустойчивости растений большое влияние оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках. В зимующих растениях в цитоплазме накапливаются сахара, а содержание крахмала снижается. Влияние сахаров на повышение морозоустойчивости растений многосторонне. Накопление Сахаров предохраняет от замерзания большой объем внутриклеточной воды, заметно уменьшает количество образующегося льда.
Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условий среды в соответствии с генотипом растения, связано с резким снижением темпов роста, переходом растения в состояние покоя.
Жизненный цикл развития озимых, двулетних и многолетних растений контролируется сезонным ритмом светового и температурного периодов. В отличие от яровых однолетних растений они начинают готовиться к перенесению неблагоприятных зимних условий с момента остановки роста и затем в течение осени во время наступления пониженных температур.
Зимостойкость растений
Зимостойкость как устойчивость к комплексу неблагоприятных факторов перезимовки.
Непосредственное действие мороза на клетки - не единственная опасность, угрожающая многолетним травянистым и древесным культурам, озимым растениям в течение зимы. Помимо прямого действия мороза растения подвергаются еще ряду неблагоприятных факторов. В течение зимы температура может существенно колебаться. Морозы нередко сменяются кратковременными и длительными оттепелями. В зимнее время нередки снежные бури, а в бесснежные зимы в более южных районах страны - и суховеи. Все это истощает растения, которые после перезимовки выходят сильно ослабленными и в последующем могут погибнуть.
Особенно многочисленные неблагоприятные воздействия испытывают травянистые многолетние и однолетние растения. На территории России в неблагоприятные годы гибель посевов озимых зерновых достигает 30-60 %. Погибают не только озимые хлеба, но и многолетние травы, плодовые и ягодные многолетние насаждения. Кроме низких температур озимые растения повреждается и гибнут от ряда других неблагоприятных факторов в зимнее время и ранней весной: выпревания, вымокания, ледяной корки, выпирания, повреждения от зимней засухи.
Выпревание, вымокание, гибель под ледяной коркой, выпирание, повреждение от зимней засухи.
Выпревание. Среди перечисленных невзгод первое место занимает выпревание растений. Гибель растений от выпревания наблюдается преимущественно в теплые зимы с большим снеговым покровом, который лежит 2-3 месяца, особенно если снег выпадает на мокрую и талую землю. Исследования показали, что причина гибели озимых от выпревания - истощение растений. Находясь под снегом при температуре около О °С в сильно увлажненной среде, почти полной темноте, т. е. в условиях, при которых процесс дыхания идет достаточно интенсивно, а фотосинтез исключен, растения постепенно расходуют сахара и другие запасы питательных веществ, накопленные в период прохождения первой фазы закаливания, и погибают от истощения (содержание Сахаров в тканях уменьшается с 20 до 2-4 %) и весенних заморозков. Такие растения весной легко повреждаются снежной плесенью, что также приводит к их гибели.
Вымокание. Вымокание проявляется преимущественно весной в пониженных местах в период таяния снега, реже во время длительных оттепелей, когда на поверхности почвы накапливается талая вода, которая не впитывается в замершую почву и может затопить растения. В этом случае причиной гибели растений служит резкий недостаток кислорода (анаэробные условия - гипоксия). У растений, оказавшихся под слоем воды, нормальное дыхание прекращается из-за недостатка кислорода в воде и почве. Отсутствие кислорода усиливает анаэробное дыхание растений, в результате чего могут образоваться токсичные вещества и растения погибают от истощения и прямого отравления организма.
Гибель под ледяной коркой. Ледяная корка образуется на полях в районах, где частые оттепели сменяются сильными морозами. Действие вымокания в этом случае может усугубляться. При этом происходит образование висячих или притертых (контактных) ледяных корок. Менее опасны висячие корки, так как они образуются сверху почвы и практически не соприкасаются с растениями; их легко разрушить катком.
При образовании же сплошной ледяной контактной корки растения полностью вмерзают в лед, что ведет к их гибели, так как и без того ослабленные от вымокания растения подвергаются очень сильному механическому давлению.
Выпирание. Повреждение и гибель растений от выпирания определяются разрывами корневой системы. Выпирание растений наблюдается, если осенью морозы наступают при отсутствии снежного покрова или если в поверхностном слое почвы мало воды (при осенней засухе), а также при оттепелях, если снеговая вода успеет всосаться в почву. В этих случаях замерзание воды начинается не с поверхности почвы, а на некоторой глубине (где есть влага). Образующаяся на глубине прослойка льда постепенно утолщается за счет продолжающегося поступления воды по почвенным капиллярам и поднимает (выпирает) верхние слои почвы вместе с растениями, что приводит к обрыву корней растений, проникших на значительную глубину.
Повреждения от зимней засухи. Устойчивый снеговой покров предохраняет озимые злаки от зимнего высыхания. Однако они в условиях бесснежной или малоснежной зимы, как и плодовые деревья и кустарники, в ряде районов России часто подвергаются опасности чрезмерного иссушения постоянными и сильными ветрами, особенно в конце зимы при значительном нагреве солнцем. Дело в том, что водный баланс растений складывается зимой крайне неблагоприятно, так как поступление воды из замерзшей почвы практически прекращается.
Для уменьшения испарения воды, неблагоприятного действия зимней засухи плодовые древесные породы образуют на ветвях мощный слой пробки, сбрасывают на зиму листья.
Яровизация
Фотопериодические реакции на сезонные изменения длины дня имеют значение для периодичности цветения многих видов как умеренных, так и тропических областей. Однако следует отметить, что среди видов умеренных широт, проявляющих фотопериодические реакции, относительно мало весеннецветущих, хотя мы постоянно сталкиваемся с тем, что значительное число «цветов цветет весной», и многие из таких весеннецветущих форм, например Ficariaverna, первоцвет (Primulavutgaris), фиалки (виды рода Viola) и т. д., проявляют выраженное сезонное поведение, оставаясь вегетативными оставшуюся часть года после обильного весеннего цветения. Можно предположить, что весеннее цветение -реакция на короткие дни зимой, но для многих видов, это, по-видимому, не так.
Конечно, длина дня не является единственным внешним фактором, изменяющимся в течение года. Ясно, что и температура также характеризуется четко выраженными сезонными изменениями, особенно в умеренных областях, хотя в отношении этого фактора наблюдаются значительные колебания, как ежедневные, так и ежегодные. Мы знаем, что сезонные изменения температуры, так же как и изменения длины дня, оказывают существенное влияние на цветение многих видов растений.
Типы растений, требующих охлаждения для перехода к цветению .
Было установлено, что многие виды, в том числе озимые однолетние, а также двулетние и многолетние травянистые растения, нуждаются в охлаждении для перехода к цветению.
Известно, что озимые однолетники и двулетники представляют собой монокарпические растения, которые требуют яровизации,- они остаются вегетативными во время первого вегетативного сезона и цветут следующей весной или ранним летом в ответ на период охлаждения, получаемый зимой. Необходимость охлаждения двулетних растений для индукции цветения была экспериментально продемонстрирована на ряде видов, таких, как свекла (Betavulgaris), сельдерей (Apiutngraveolens), капуста и другие культивируемые сорта рода Brassiса, колокольчик (Campanulamedium), лунник (Lunariabiennis), наперстянка (Digitalispurpurea) и другие. Если растения наперстянки, которые в нормальных условиях ведут себя как двулетники, т. е. зацветают на второй год после прорастания, содержать в оранжерее, они могут оставаться вегетативными несколько лет. В районах с мягкой зимой капуста в течение нескольких лет может расти в открытом грунте без «образования стрелки» (т. е. цветения) весной, что обычно происходит в районах с холодной зимой. Такие виды обязательно требуют яровизации, однако у ряда других видов цветение ускоряется при воздействии на них холодом, но может наступать и без яровизации; к таким видам, проявляющим факультативную потребность в холоде, относятся салат (Lactucasaiiva), шпинат (Spinaciaoleracea) и позднецветущие сорта гороха (Pistimsa-tivum).
Так же как и двулетние, многие многолетние виды нуждаются в воздействии холодом и не зацветают без ежегодного зимнего охлаждения. Из обычных многолетних растений в холодовом воздействии нуждаются первоцвет (Primulavulgaris), фиалки (Violaspp.), лакфиоль (Cheiranthuscheirii и С. allionii), левкой (Mathiolaincarna), некоторые сорта хризантем (Chrisant-hemummorifolium), виды рода Aster, турецкая гвоздика (Dianthus), плевел (Loliumperenne). Многолетние виды требуют переяровизации каждую зиму.
Вполне вероятно, что и у других весеннецветущих многолетников можно обнаружить потребность в охлаждении. Весенне-цветущие луковичные растения, такие, как нарциссы, гиацинты, пролески (Endymionnonscriptus), крокусы и т. д. не требуют охлаждения для заложения цветков, поскольку примордий цветка заложился в луковице предыдущим летом, но их рост в значительной степени зависит от температурных условий. Например, у тюльпана началу цветения благоприятствуют относительно высокие температуры (20°С), но для удлинения стебля и роста листьев оптимальной температурой вначале является 8-9 °С с последовательным повышением на более поздних стадиях до 13, 17 и 23°С. Аналогичные реакции на температуру характерны для гиацинтов и нарциссов.
У многих видов заложение цветка происходит не во время самого периода охлаждения и начинается лишь после того, как растение подверглось действию более высоких температур, следующих за охлаждением.
Таким образом, хотя при низких температурах метаболизм у большинства растений значительно замедляется, не вызывает сомнения, что яровизация включает активные физиологические процессы, природа которых пока еще совершенно неизвестна.
Жароустойчивость растений
Жароустойчивость (жаровыносливость) - способность растений переносить действие высоких температур, перегрев. Это генетически обусловленный признак. Виды растений различаются по выносливости к высоким температурам.
По жароустойчивости выделяют три группы растений.
Жаростойкие - термофильные сине-зеленые водоросли и бактерии горячих минеральных источников, способные переносить повышение температуры до 75-100 °С. Жароустойчивость термофильных микроорганизмов определяется высоким уровнем метаболизма, повышенным содержанием РНК в клетках, устойчивостью белка цитоплазмы к тепловой коагуляции.
Жаровыносливые - растения пустынь и сухих мест обитания (суккуленты, некоторые кактусы, представители семейства Толстянковые), выдерживающие нагревание солнечными лучами до 50-65єС. Жароустойчивость суккулентов во многом определяется повышенными вязкостью цитоплазмы и содержанием связанной воды в клетках, пониженным обменом веществ.
Нежаростойкие - мезофитные и водные растения. Мезофиты открытых мест переносят кратковременное действие температур 40-47 єС, затененных мест - около 40-42 °С, водные растения выдерживают повышение температуры до 38-42 °С. Из сельскохозяйственных наиболее жаровыносливы теплолюбивые растения южных широт (сорго, рис, хлопчатник, клещевина и др.).
Многие мезофиты переносят высокую температуру воздуха и избегают перегрева благодаря интенсивной транспирации, снижающей температуру листьев. Более жаростойкие мезофиты отличаются повышенной вязкостью цитоплазмы и усиленным синтезом жаростойких белков-ферментов.
Растения выработали систему морфологических и физиологических приспособлений, защищающих их от тепловых повреждений: светлую окраску поверхности, отражающую инсоляцию; складывание и скручивание листьев; опушения или чешуйки, защищающие от перегрева глубжележащие ткани; тонкие слои пробковой ткани, предохраняющие флоэму и камбий; большую толщину кутикулярного слоя; высокое содержание углеводов и малое - воды в цитоплазме и др.
На тепловой стресс растения очень быстро реагируют индуктивной адаптацией. К воздействию высоких температур они могут подготовиться за несколько часов. Так, в жаркие дни устойчивость растений к высоким температурам после полудня выше, чем утром. Обычно эта устойчивость временная, она не закрепляется и довольно быстро исчезает, если становится прохладно. Обратимость теплового воздействия может составлять от нескольких часов до 20 дней. В период образования генеративных органов жаростойкость однолетних и двулетних растений снижается.
Засухоустойчивость растений
Обычным явлением для многих регионов России и государств СНГ стали засухи. Засуха - это длительный бездождливый период, сопровождаемый снижением относительной влажности воздуха, влажности почвы и повышением температуры, когда не обеспечиваются нормальные потребности растений в воде. На территории России имеются регионы неустойчивого увлажнения с годовым количеством осадков 250-500 мм и засушливые, с количеством осадков менее 250 мм в год при испаряемости более 1000 мм.
Засухоустойчивость - способность растений переносить длительные засушливые периоды, значительный водный дефицит, обезвоживание клеток, тканей и органов. При этом ущерб урожая зависит от продолжительности засухи и ее напряженности. Различают засуху почвенную и атмосферную.
Почвенная засуха вызывается длительным отсутствием дождей в сочетании с высокой температурой воздуха и солнечной инсоляцией, повышенным испарением с поверхности почвы и транспирацией, сильными ветрами. Все это приводит к иссушению корнеобитаемого слоя почвы, снижению запаса доступной для растений воды при пониженной влажности воздуха. Атмосферная засуха характеризуется высокой температурой и низкой относительной влажностью воздуха (10-20 %). Жесткая атмосферная засуха вызывается перемещением масс сухого и горячего воздуха - суховея. К тяжелым последствиям приводит мгла, когда суховей сопровождается появлением в воздухе почвенных частиц (пыльные бури).
Атмосферная засуха, резко усиливая испарение воды с поверхности почвы и транспирацию, способствует нарушению согласованности скоростей поступления из почвы в надземные органы воды и потери ее растением, в результате растение завядает. Однако при хорошем развитии корневой системы атмосферная засуха не причиняет растениям большого вреда, если температура не превышает переносимый растениями предел. Продолжительная атмосферная засуха в отсутствие дождей приводит к почвенной засухе, которая более опасна для растений.
Засухоустойчивость обусловлена генетически определенной приспособленностью растений к условиям места обитания, а также адаптацией к недостатку воды. Засухоустойчивость выражается в способности растений переносить значительное обезвоживание за счет развития высокого водного потенциала тканей при функциональной сохранности клеточных структур, а также за счет адаптивных морфологических особенностей стебля, листьев, генеративных органов, повышающих их выносливость, толерантность к действию длительной засухи.
Типы растений по отношению к водному режиму
Растения засушливых областей называются ксерофитами (от греческого хеrоs - сухой). Они способны в процессе индивидуального развития приспосабливаться к атмосферной и почвенной засухе. Характерные признаки ксерофитов - незначительные размеры их испаряющей поверхности, а также небольшие размеры надземной части по сравнению с подземной. Ксерофиты - это обычно травы или низкорослые кустарники. Их делят на несколько типов. Приводим классификацию ксерофитов по П. А. Генкелю.
Суккуленты - очень стойкие к перегреву и устойчивые к обезвоживанию, во время засухи они не испытывают недостатка воды, потому что содержат большое количество ее и медленно расходуют. Корневая система у них разветвлена во все стороны в верхних слоях почвы, благодаря чему в дождливые периоды растения быстро всасывают воду. Это кактусы, алоэ, очиток, молодило.
Эвксерофиты - жаростойкие растения, которые хорошо переносят засуху. К этой группе относятся такие степные растения, как вероника сизая, астра мохнатая, полынь голубая, арбуз колоцинт, верблюжья колючка и др. У них незначительная транспирация, высокое осмотическое давление, цитоплазма отличается высокой эластичностью и вязкостью, корневая система очень разветвлена, и основная ее масса размещена в верхнем слое почвы (50-60 см). Эти ксерофиты способны сбрасывать листья и даже целые ветви.
Гемиксерофиты, или полуксерофиты - это растения, которые неспособны переносить обезвоживание и перегрев. Вязкость и эластичность протопласта у них незначительная, отличается высокой транспирацией, глубокой корневой системой, которая может достигать подпочвенной воды, что обеспечивает бесперебойное снабжение растения водой. К этой группе относятся шалфей, резак обычный и др.
Стипаксерофшпы - это ковыль, тырса и другие узколистные степные злаки. Они устойчивы к перегреву, хорошо используют влагу кратковременных дождей. Выдерживают лишь кратковременную нехватку воды в почве.
Пойкилоксерофиты - растения, не регулирующие своего водного режима. Это в основном лишайники, которые могут высыхать до воздушно-сухого состояния и снова проявлять жизнедеятельность после дождей.
Гигрофиты (от греческого hihros - влажный). У растений, относящихся к этой группе, нет приспособлений, ограничивающих расход воды. Для гигрофитов характерны сравнительно большие размеры клеток, тонкостенная оболочка, слабоодревесневшие стенки сосудов, древесных и лубяных волокон, тонкая кутикула и малоутолщенные внешние стенки эпидермиса, большие устьица и незначительное количество их на единицу поверхности, большая листовая пластинка, плохо развитые механические ткани, редкая сеть жилок в листе, большая кутикулярная транспирация, длинный стебель, недостаточно развитая корневая система. По строению гигрофиты приближаются к теневыносливым растениям, но имеют своеобразную гигроморфную структуру. Незначительный недостаток воды в почве вызывает быстрое завядание гигрофитов. Осмотическое давление клеточного сока в них невысокое. К ним относятся манник, багульник, брусника, лох.
По условиям произрастания и особенностям строения к гигрофитам очень близки растения с частично или полностью погруженными в воду или плавающими на ее поверхности листьями, которые называются гидрофитами.
Мезофиты (от греческого mesos - средний, промежуточный). Растения этой экологической группы произрастают в условиях достаточного увлажнения. Осмотическое давление клеточного сока у мезофитов 1-1,5 тыс. кПа. Они легко завядают. К мезофитам относятся большинство луговых злаков и бобовых -пырей ползучий, лисохвост луговой, тимофеевка луговая, люцерна синяя и др. Из полевых культур твердые и мягкие пшеницы, кукуруза, овес, горох, соя, сахарная свекла, конопля, почти все плодовые (за исключением миндаля, винограда), многие овощные культуры (морковь, помидорыи др.).
Транспирирующие органы – листья отличаются значительной пластичностью; в зависимости от условий произрастания в их строении наблюдаются довольно большие отличия. Даже листья одного растения при разном водоснабжении и освещении имеют различия в строении. Установлены определенные закономерности в строении листьев в зависимости от расположения их на растении.
В. Р. Заленский обнаружил изменения в анатомическом строении листьев по ярусам. Он установил, что у листьев верхнего яруса наблюдаются закономерные изменения в сторону усиления ксероморфизма, т. е. происходит образование структур, повышающих засухоустойчивость этих листьев. Листья, расположенные в верхней части стебля, всегда отличаются от нижних, а именно: чем выше расположен лист на стебле, тем меньше размеры его клеток, большее количество устьиц и меньше их размеры, большее количество волосков на единицу поверхности, гуще сеть проводящих пучков, сильнее развита палисадная ткань. Все эти признаки характеризуют ксерофилию, т. е. образование структур, способствующих повышению засухоустойчивости.
С определенной анатомической структурой связаны и физиологические особенности, а именно: верхние листья отличаются более высокой ассимиляционной способностью и более интенсивной транспирацией. Концентрация сока в верхних листьях также более высокая, в связи с чем может происходить оттягивание воды верхними листьями от нижних, засыхание и отмирание нижних листьев. Структура органов и тканей, повышающая засухоустойчивость растений, называется ксероморфизмом. Отличительные особенности в структуре листьев верхнего яруса объясняются тем, что они развиваются в условиях несколько затрудненного водоснабжения.
Для уравнения баланса между поступлением и расходом воды в растении образовалась сложная система анатомо-физиологических приспособлений. Такие приспособления наблюдаются у ксерофитов, гигрофитов, мезофитов.
Результаты исследований показали, что приспособительные свойства у засухоустойчивых форм растений возникают под влиянием условий их существования.
ЗАКЛЮЧЕНИЕ
Удивительная гармония живой природы, ее совершенство создаются самой природой: борьбой за выживание. Формы приспособлений у растений и животных бесконечно разнообразны. Весь животный и растительный мир со времени своего появления совершенствуется по пути целесообразных приспособлений к условиям обитания: к воде, к воздуху, солнечному свету, силы тяжести и т.д.
ЛИТЕРАТУРА
1. Володько И.К. ""Микроэлементы и устойчивость растений к неблагоприятным условиям"", Минск, Наука и техника, 1983г.
2. Горышина Т.К. ""Экология растений"", уч. Пособие для ВУЗов, Москва, В. школа, 1979г.
3. Прокофьев А.А. ""Проблемы засухоустойчивости растений"", Москва, Наука, 1978г.
4. Сергеева К.А. ""Физиологические и биохимические основы зимостойкости древесных растений"", Москва, Наука, 1971г
5. Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
Приспособленность онтогенеза растений к условиям среды является результатом их эволюционного развития (изменчивости, наследственности, отбора). На протяжении филогенеза каждого вида растений в процессе эволюции выработались определенные потребности индивидуума к условиям существования и приспособленность к занимаемой им экологической нише. Влаголюбие и теневыносливость, жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в ходе эволюции в результате длительного действия соответствующих условий. Так, теплолюбивые растения и растения короткого дня характерны для южных широт, менее требовательные к теплу и растения длинного дня - для северных.
В природе в одном географическом регионе каждый вид растений занимает экологическую нишу, соответствующую его биологическим особенностям: влаголюбивые - ближе к водоемам, теневыносливые - под пологом леса и т. д. Наследственность растений формируется под влиянием определенных условий внешней среды. Важное значение имеют и внешние условия онтогенеза растений.
В большинстве случаев растения и посевы (посадки) сельскохозяйственных культур, испытывая действие тех или иных неблагоприятных факторов, проявляют устойчивость к ним как результат приспособления к условиям существования, сложившимся исторически, что отмечал еще К. А. Тимирязев.
1. Основные среды жизни.
При изучении окружающей среды (среды обитания растений и животных и производственной деятельности человека) выделяют следующие ее основные составляющие: воздушную среду; водную среду (гидросферу); животный мир(человек, домашние и дикие животные, в том числе рыбы и птицы); растительный мир (культурные и дикие растения в том числе растущие в воде);почву(растительный слой);недра(верхняя часть земной коры, в пределах которой возможна добыча полезных ископаемых); климатическую и акустическую среду.
Воздушная среда может быть наружной, в которой большинство людей проводят меньшую часть времени (до 10-15%), внутренней производственной (в ней человек проводит до 25-30% своего времени) и внутренней жилой, где люди пребывают большую часть времени (до 60-70% и более).
Наружный воздух у поверхности земли содержит по объему: 78,08% азота; 20,95% кислорода; 0,94% инертных газов и 0,03% углекислого газа. На высоте 5 км содержание кислорода остается тем же, а азота увеличивается до 78,89%. Часто воздух у поверхности земли имеет различные примеси, особенно в городах: там он содержит более 40 ингредиентов, чуждых природной воздушной среде. Внутренний воздух в жилищах, как правило, имеет
повышенное содержание углекислого газа, а внутренний воздух производственных помещений обычно содержит примеси, характер которых определяется технологией производства. Среди газов выделяется водяной пар, который попадает в атмосферу в результате испарений с Земли. Большая его часть (90%) сосредоточена в самом нижнем пятикилометровом слое атмосферы, с высотой его количество очень быстро уменьшается. Атмосфера содержит много пыли, которая попадает туда с поверхности Земли и частично из космоса. При сильных волнениях ветры подхватывают водяные брызги из морей и океанов. Так попадают в атмосферу из воды частицы соли. В результате извержения вулканов, лесных пожаров, работы промышленных объектов и т.д. воздух загрязняется продуктами неполного сгорания. Больше всего пыли и других примесей в приземном слое воздуха. Даже после дождя в 1 см содержится около 30 тыс. пылинок, а в сухую погоду их в сухую погоду их в несколько раз больше.
Все эти мельчайшие примеси влияют на цвет неба. Молекулы газов рассеивают коротковолновую часть спектра солнечного луча, т.е. фиолетовые и синие лучи. Поэтому днем небо голубого цвета. А частицы примесей, которые значительно крупней молекул газов, рассеивают световые лучи почти всех длин волн. Поэтому, когда воздух запылен или в нем содержатся капельки воды, небо становится белесоватым. На больших высотах небо темно-фиолетовое и даже черное.
В результате происходящего на Земле фотосинтеза растительность ежегодно образует 100 млрд. т. органических веществ (около половины приходится на долю морей и океанов), усваивая при этом около 200 млрд. т. углекислого газа и выделяя во внешнюю среду около 145 млрд.т. свободного кислорода, полагают, что благодаря фотосинтезу образуется весь кислород атмосферы. О роли в этом круговороте зеленых насаждений говорят следующие данные: 1 га зеленых насаждений в среднем за 1 час очищает воздух от 8 кг углекислого газа (выделяемого за это время при дыхании 200 человек). Взрослое дерево за сутки выделяет 180 литров кислорода, а за пять месяцев (с мая по сентябрь) оно поглощает около 44 кг углекислого газа.
Количество выделяемого кислорода и поглощаемого углекислого газа зависит от возраста зеленых насаждений, видового состава, плотности посадки и других факторов.
Не меньшее значение имеют и морские растения - фитопланктон(в основном водоросли и бактерии), высвобождающие путем фотосинтеза кислород.
Водная среда включает поверхностные и подземные воды. Поверхностные воды в основном сосредоточены в океане, содержанием 1 млрд. 375 млн. кубических километров - около 98% всей воды на Земле. Поверхность океана (акватория) составляет 361 млн. квадратных километров. Она примерно в 2,4 раза больше площади суши--территории, занимающей 149 млн. квадратных километров. Вода в океане соленая, причем большая ее часть (более 1 млрд. кубических километров) сохраняет постоянную соленость около 3,5% и температуру, примерно равную 3,7є С. Заметные различия в солености и температуре наблюдаются почти исключительно в поверхностном слое воды, а также в окраинных и особенно в средиземных морях. Содержание растворенного кислорода в воде существенно уменьшается на глубине 50-60 метров.
Подземные воды бывают солеными, солоноватыми (меньшей солености) и пресными; существующие геотермальные воды имеют повышенную температуру (более 30єС).
Для производственной деятельности человечества и его хозяйственно-бытовых нужд требуется пресная вода, количество которой составляет всего лишь 2,7% общего объема воды на Земле, причем очень малая ее доля (всего 0,36%) имеется в легкодоступных для добычи местах. Большая часть пресной воды содержится в снегах и пресноводных айсбергах, находящихся в районах в основном Южного полярного круга.
Годовой мировой речной сток пресной воды составляет 37,3 тыс. кубических километров. Кроме того, может использоваться часть подземных вод, равная 13 тыс. кубическим километрам. К сожалению, большая часть речного стока в России, составляющая около 5000 кубических километров, приходится на малоплодородные и малозаселенные северные территории.
Климатическая среда является важным фактором, определяющим развитие различных видов животного, растительного мира и его плодородие. Характерной особенностью России является то, что большая часть ее территории имеет значительно более холодный климат, чем в других странах.
Все рассмотренные составляющие окружающей среды входят в
БИОСФЕРУ: оболочку Земли, включающую часть атмосферы, гидросферу и верхнюю часть литосферы, которые взаимно связанны сложными биохимическими циклами миграции вещества и энергии, геологическую оболочку Земли, населенную живыми организмами. Верхний предел жизни биосферы ограничен интенсивной концентрацией ультрафиолетовых лучей; нижний - высокой температурой земных недр (свыше100`С). Крайних пределов ее достигают только низшие организмы - бактерии.
Адаптация (приспособление) растения к конкретным условиям среды обеспечивается за счет физиологических механизмов (физиологическая адаптация), а у популяции организмов (вида) - благодаря механизмам генетической изменчивости, наследственности и отбора (генетическая адаптация). Факторы внешней среды могут изменяться закономерно и случайно. Закономерно изменяющиеся условия среды (смена сезонов года) вырабатывают у растений генетическую приспособленность к этим условиям.
В естественных для вида природных условиях произрастания или возделывания растения в процессе своего роста и развития часто испытывают воздействие неблагоприятных факторов внешней среды, к которым относят температурные колебания, засуху, избыточное увлажнение, засоленность почвы и т. д. Каждое растение обладает способностью к адаптации в меняющихся условиях внешней среды в пределах, обусловленных его генотипом. Чем выше способность растения изменять метаболизм в соответствии с окружающей средой, тем шире норма реакции данного растения и лучше способность к адаптации. Это свойство отличает устойчивые сорта сельскохозяйственных культур. Как правило, несильные и кратковременные изменения факторов внешней среды не приводят к существенным нарушениям физиологических функций растений, что обусловлено их способностью сохранять относительно стабильное состояние при изменяющихся условиях внешней среды, т. е. поддерживать гомеостаз. Однако резкие и длительные воздействия приводят к нарушению многих функций растения, а часто и к его гибели.
При действии неблагоприятных условий снижение физиологических процессов и функций может достигать критических уровней, не обеспечивающих реализацию генетической программы онтогенеза, нарушаются энергетический обмен, системы регуляции, белковый обмен и другие жизненно важные функции растительного организма. При воздействии на растение неблагоприятных факторов (стрессоров) в нем возникает напряженное состояние, отклонение от нормы - стресс. Стресс - общая неспецифическая адаптационная реакция организма на действие любых неблагоприятных факторов. Выделяют три основные группы факторов, вызывающих стресс у растений: физические - недостаточная или избыточная влажность, освещенность, температура, радиоактивное излучение, механические воздействия; химические - соли, газы, ксенобиотики (гербициды, инсектициды, фунгициды, промышленные отходы и др.); биологические - поражение возбудителями болезней или вредителями, конкуренция е другими растениями, влияние животных, цветение, созревание плодов.
Реакции на неблагоприятные факторы среды только при некоторых условиях являются губительными для живых организмов, а в большинстве случаев они имеют адаптивное значение. Поэтому эти ответные реакции были названы Селье «общим адаптационным синдромом». В более поздних работах термины «стресс» и «общий адаптационный синдром» он употреблял как синонимы.
Адаптация — это генетически детерминированный процесс формирования защитных систем, которые обеспечивают повышение устойчивости и протекание онтогенеза в неблагоприятных для него условиях.
Адаптация является одним из важнейших механизмов, который повышает устойчивость биологической системы, в том числе растительного организма, в изменившихся условиях существования. Чем лучше организм адаптирован к какому-то фактору, тем он устойчивее к его колебаниям.
Генотипически обусловленная способность организма изменять метаболизм в определенных пределах в зависимости от действия внешней среды называется нормой реакции . Она контролируется генотипом и свойственна всем живым организмам. Большинство модификаций, которые возникают в пределах нормы реакции, имеют адаптивное значение. Они соответствуют изменениям среды обитания и обеспечивают лучшую выживаемость растений при колебаниях условии окружающей среды. В этой связи такие модификации имеют эволюционное значение. Термин «норма реакции» введен В.Л. Йогансеном (1909).
Чем больше способность вида или сорта модифицироваться в соответствии с окружающей средой, тем шире его норма реакции и выше способность к адаптации. Это свойство отличает устойчивые сорта сельскохозяйственных культур. Как правило, несильные и кратковременные изменения факторов внешней среды не приводят к существенным нарушениям физиологических функций растений. Это обусловлено их способностью сохранять относительное динамическое равновесие внутренней среды и устойчивость основных физиологических функций в условиях изменяющейся внешней среды. В то же время резкие и продолжительные воздействия приводят к нарушению многих функций растения, а нередко и к его гибели.
Адаптация включает в себя все процессы и приспособления (анатомические, морфологические, физиологические, поведенческие и др.), которые способствуют повышению устойчивости и способствуют выживанию вида.
1. Анатомо-морфологические приспособления . У некоторых представителей ксерофитов длина корневой системы достигает несколько десятков метров, что позволяет растению использовать грунтовую воду и не испытывать недостатка влаги в условиях почвенной и атмосферной засухи. У других ксерофитов наличие толстой кутикулы, опушенность листьев, превращение листьев в колючки уменьшают потери воды, что очень важно в условиях недостатка влаги.
Жгучие волоски и колючки защищают растения от поедания животными.
Деревья в тундре или на больших горных высотах имеют вид приземистых стелющихся кустарников, зимой они засыпаются снегом, который защищает их от сильных морозов.
В горных районах с большими суточными колебаниями температуры растения часто имеют форму распластанных подушек с плотно расположенными многочисленными стеблями. Это позволяет сохранять внутри подушек влагу и относительно равномерную в течение суток температуру.
У болотных и водных растений формируется специальная воздухоносная паренхима (аэренхима), которая является резервуаром воздуха и облегчает дыхание частей растения, погруженных в воду.
2. Физиолого-биохимические приспособления . У суккулентов приспособлением для произрастания в условиях пустынь и полупустынь является усвоение СО 2 в ходе фотосинтеза по CAM-пути. У этих растений устьица днем закрыты. Таким образом, растение сохраняет внутренние запасы воды от испарения. В пустынях вода является главным фактором, ограничивающим рост растений. Устьица открываются ночью, и в это время происходит поступление СО 2 в фотосинтезирующие ткани. Последующее вовлечение СО 2 в фотосинтетический цикл происходит днем уже при закрытых устьицах.
К физиолого-биохимическим приспособлениям относятся способность устьиц открываться и закрываться, в зависимости от внешних условий. Синтез в клетках абсцизовой кислоты, пролина, защитных белков, фитоалексинов, фитонцидов, повышение активности ферментов, противодействующих окислительному распаду органических веществ, накопление в клетках сахаров и ряд других изменений в обмене веществ содействует повышению устойчивости растений к неблагоприятным условиям внешней среды.
Одна и та же биохимическая реакция может осуществляться несколькими молекулярными формами одного и того же фермента (изоферментами), при этом каждая изоформа проявляет каталитическую активность в относительно узком диапазоне некоторого параметра окружающей среды, например температуры. Наличие целого ряда изоферментов позволяет растению осуществлять реакцию в значительно более широком диапазоне температур, по сравнению с каждым отдельным изоферментом. Это дает возможность растению успешно выполнять жизненные функции в изменяющихся температурных условиях.
3. Поведенческие приспособления, или избегание действия неблагоприятного фактора . Примером могут служить эфемеры и эфемероиды (мак, звездчатка, крокусы, тюльпаны, подснежники). Они проходят весь цикл своего развития весной за 1,5-2 месяца, еще до наступления жары и засухи. Таким образом, они как бы уходят, или избегают попадания под влияние стрессора. Подобным образом раннеспелые сорта сельскохозяйственных культур формируют урожай до наступления неблагоприятных сезонных явлений: августовских туманов, дождей, заморозков. Поэтому селекция многих сельскохозяйственных культур направлена на создание раннеспелых сортов. Многолетние растения зимуют в виде корневищ и луковиц в почве под снегом, защищающим их от вымерзания.
Адаптация растений к неблагоприятным факторам осуществляется одновременно на многих уровнях регуляции — от отдельной клетки до фитоценоза. Чем выше уровень организации (клетка организм, популяция) тем большее число механизмов одновременно участвует в адаптации растений к стрессам.
Регуляция метаболических и адаптационных процессов внутри клетки осуществляется с помощью систем: метаболической (ферментативной); генетической; мембранной. Эти системы тесно связаны между собой. Так, свойства мембран зависят от генной активности, а дифференциальная активность самих генов находится под контролем мембран. Синтез ферментов и их активность контролируются на генетическом уровне, в то же время ферменты регулируют нуклеиновый обмен в клетке.
На организменном уровне к клеточным механизмам адаптации добавляются новые, отражающие взаимодействие органов. В неблагоприятных условиях растения создают и сохраняют такое количество плодоэлементов, которое в достаточном количестве обеспечено необходимыми веществами, чтобы сформировать полноценные семена. Например, в соцветиях культурных злаков и в кронах плодовых деревьев в неблагоприятных условиях более половины заложившихся завязей могут опасть. Такие изменения основаны на конкурентных отношениях между органами за физиологически активные и питательные вещества.
В условиях стрессов резко ускоряются процессы старения и опадения нижних листьев. При этом нужные растениям вещества перемещаются из них в молодые органы, отвечая стратегии выживания организма. Благодаря реутилизации питательных веществ из нижних листьев сохраняются жизнеспособными более молодые — верхние листья.
Действуют механизмы регенерации утраченных органов. Например, поверхность ранения покрывается вторичной покровной тканью (раневой перидермой), рана на стволе или ветке зарубцовывается наплывами (каллюсами). При утрате верхушечного побега у растений пробуждаются спящие почки и усиленно развиваются боковые побеги. Весеннее восстановление листьев вместо опавших осенью — это также пример естественной регенерации органов. Регенерация как биологическое приспособление, обеспечивающее вегетативное размножение растений отрезками корня, корневища, слоевища, стеблевыми и листовыми черенками, изолированными клетками, отдельными протопластами, имеет большое практическое значение для растениеводства, плодоводства, лесоводства, декоративного садоводства и пр.
В процессах защиты и адаптации на уровне растения участвует и гормональная система. Например, при действии неблагоприятных условий в растении резко возрастает содержание ингибиторов роста: этилена и абсциссой кислоты. Они снижают обмен веществ, тормозят ростовые процессы, ускоряют старение, опадение органов, переход растения в состояние покоя. Торможение функциональной активности в условиях стресса под влиянием ингибиторов роста является характерной для растений реакцией. Одновременно с этим в тканях снижается содержание стимуляторов роста: цитокинина, ауксина и гиббереллинов.
На популяционном уровне присоединяется отбор, который приводит к появлению более приспособленных организмов. Возможность отбора определяется существованием внутрипопуляционной изменчивости устойчивости растений к разным факторам внешней среды. Примером внутрипопуляционной изменчивости по устойчивости может служить недружность появления всходов на засоленной почве и увеличение варьирования сроков прорастания при усилении действия стрессора.
Вид в современном представлении состоит из большого числа биотипов — более мелких экологических единиц, генетически одинаковых, но проявляющих разную устойчивость к факторам внешней среды. В различных условиях не все биотипы одинаково жизненны, и в результате конкуренции остаются лишь те из них, которые наиболее отвечают данным условиям. То есть, устойчивость популяции (сорта) к тому или иному фактору определяется устойчивостью составляющих популяцию организмов. Устойчивые сорта имеют в своем составе набор биотипов, обеспечивающих хорошую продуктивность даже в неблагоприятных условиях.
Вместе с тем, в процессе многолетнего культивирования у сортов изменяется состав и соотношение биотипов в популяции, что отражается на продуктивности и качестве сорта, часто не в лучшую сторону.
Итак, адаптация включает в себя все процессы и приспособления, повышающие устойчивость растений к неблагоприятным условиям среды (анатомические, морфологические, физиологические, биохимические, поведенческие, популяционные и др.)
Но для выбора наиболее эффективного пути адаптации главным является время, в течение которого организм должен приспособиться к новым условиям.
При внезапном действии экстремального фактора ответ не может быть отложен, он должен последовать незамедлительно, чтобы исключить необратимые повреждения растения. При длительных воздействиях небольшой силы адаптационные перестройки происходят постепенно, при этом увеличивается выбор возможных стратегий.
В этой связи различают три главные стратегии адаптации: эволюционные , онтогенетические и срочные . Задача стратегии — эффективное использование имеющихся ресурсов для достижения основной цели — выживания организма в условиях стресса. Стратегия адаптации направлена на поддержание структурной целостности жизненно важных макромолекул и функциональной активности клеточных структур, сохранение систем регуляции жизнедеятельности, обеспечение растений энергией.
Эволюционные, или филогенетические адаптации (филогенез — развитие биологического вида во времени) — это адаптации, возникающие в ходе эволюционного процесса на основе генетических мутаций, отбора и передающиеся по наследству. Они являются наиболее надежными для выживания растений.
У каждого вида растений в процессе эволюции выработались определенные потребности к условиям существования и приспособленность к занимаемой им экологической нише, стойкое приспособление организма к среде обитания. Влаголюбие и теневыносливость, жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в результате длительного действия соответствующих условий. Так, теплолюбивые и короткодневные растения характерны для южных широт, менее требовательные к теплу и длиннодневные растения — для северных. Хорошо известны многочисленные эволюционные адаптации к засухе растений-ксерофитов: экономное расходование воды, глубоко залегающая корневая система, сбрасывание листьев и переход в состояние покоя и другие приспособления.
В этой связи сорта сельскохозяйственных растений проявляют устойчивость именно к тем факторам внешней среды, на фоне которых проводится селекция и отбор продуктивных форм. Если отбор проходит в ряде последовательных генераций на фоне постоянного влияния какого-либо неблагоприятного фактора, то устойчивость сорта к нему может быть существенно увеличена. Закономерно, что сорта селекции НИИ сельского хозяйства Юго-Востока (г. Саратов), более устойчивы к засухе, чем сорта, созданные в селекционных центрах Московской области. Таким же путем в экологических зонах с неблагоприятными почвенноклиматическими условиями сформировались устойчивые местные сорта растений, а эндемичные виды растений устойчивы именно к тому стрессору, который выражен в ареале их обитания.
Характеристика устойчивости сортов яровой пшеницы из коллекции Всероссийского института растениеводства (Семенов и др., 2005)
| Сорт | Происхождение | Устойчивость |
| Энита | Подмосковье | Средне засухоустойчивый |
| Саратовская 29 | Саратовская обл. | Засухоустойчивый |
| Комета | Свердловская обл. | Засухоустойчивый |
| Каразино | Бразилия | Кислотоустойчивый |
| Прелюдия | Бразилия | Кислотоустойчивый |
| Колониас | Бразилия | Кислотоустойчивый |
| Тринтани | Бразилия | Кислотоустойчивый |
| ППГ-56 | Казахстан | Солеустойчивый |
| Ошская | Киргизия | Солеустойчивый |
| Сурхак 5688 | Таджикистан | Солеустойчивый |
| Мессель | Норвегия | Соленеустойчивый |
В природной обстановке условия среды обычно изменяются очень быстро, и времени, в течение которого стрессовый фактор достигает повреждающего уровня, недостаточно для формирования эволюционных приспособлений. В этих случаях растения используют не постоянные, а индуцируемые стрессором защитные механизмы, формирование которых генетически предопределено (детерминировано).
Онтогенетические (фенотипические) адаптации не связаны с генетическими мутациями и не передаются по наследству. Формирование такого рода адаптаций требует сравнительно много времени, поэтому их называют долговременными адаптациями. Одним из таких механизмов является способность ряда растений формировать водосберегающий путь фотосинтеза CAM-типа в условиях водного дефицита, вызванного засухой, засолением, действием низких температур и других стрессорами.
Эта адаптация связана с индукцией экспрессии «неактивного» в нормальных условиях гена фосфоенолпируваткарбоксилазы и генов других ферментов CAM-пути усвоения СО 2 , с биосинтезом осмолитов (пролина), с активацией антиоксидантных систем и изменением суточных ритмов устьичных движений. Все это приводит к очень экономному расходованию воды.
У полевых культур, например, у кукурузы, аэренхима в обычных условиях произрастания отсутствует. Но в условиях затопления и недостатка в тканях кислорода в корнях у нее происходит гибель части клеток первичной коры корня и стебля (апоптоз, или программируемая смерть клеток). На их месте образуются полости, по которым кислород из надземной части растения транспортируется в корневую систему. Сигналом для гибели клеток является синтез этилена.
Срочная адаптация происходит при быстрых и интенсивных изменениях условий обитания. В основе ее лежит образование и функционирование шоковых защитных систем. К шоковым защитным системам относятся, например, система белков теплового шока, которая образуется в ответ на быстрое повышение температуры. Эти механизмы обеспечивают кратковременные условия выживания при действии повреждающего фактора и тем самым создают предпосылки для формирования более надежных долговременных специализированных механизмов адаптации. Примером специализированных механизмов адаптации является новообразование антифризных белков при низких температурах или синтез сахаров в процессе перезимовки озимых культур. Вместе с тем, если повреждающее действие фактора превышает защитные и репарационные возможности организма, то неминуемо наступает смерть. В этом случае организм погибает на этапе срочной или на этапе специализированной адаптации в зависимости от интенсивности и продолжительности действия экстремального фактора.
Различают специфические и неспецифические (общие) ответные реакции растений на стрессор.
Неспецифические реакции не зависят от природы действующего фактора. Они одни и те же при действии высокой и низкой температуры, недостатка или избытка влаги, высокой концентрации солей в почве или вредных газов в воздухе. Во всех случаях в клетках растений повышается проницаемость мембран, нарушается дыхание, возрастает гидролитический распад веществ, увеличивается синтез этилена и абсцизовой кислоты, тормозится деление и растяжение клеток.
В таблице представлен комплекс неспецифических изменений, протекающих у растений под влиянием различных факторов внешней среды.
Изменение физиологических параметров у растений под действием стрессовых условий (по Г.В, Удовенко, 1995)
| Параметры | Характер изменения параметров в условиях | |||
| засухи | засоления | высокой температуры | низкой температуры | |
| Концентрация ионов в тканях | Растет | Растет | Растет | Растет |
| Активность воды в клетке | Падает | Падает | Падает | Падает |
| Осмотический потенциал клетки | Растет | Растет | Растет | Растет |
| Водоудерживающая способность | Растет | Растет | Растет | — |
| Водный дефицит | Растет | Растет | Растет | — |
| Проницаемость протоплазмы | Растет | Растет | Растет | — |
| Интенсивность транспирации | Падает | Падает | Растет | Падает |
| Эффективность транспирации | Падает | Падает | Падает | Падает |
| Энергетическая эффективность дыхания | Падает | Падает | Падает | — |
| Интенсивность дыхания | Растет | Растет | Растет | — |
| Фотофосфорилирование | Снижается | Снижается | — | Снижается |
| Стабилизация ядерной ДНК | Растет | Растет | Растет | Растет |
| Функциональнаяя активность ДНК | Снижается | Снижается | Снижается | Снижается |
| Концентрация пролина | Растет | Растет | Растет | — |
| Содержание водорастворимых белков | Растет | Растет | Растет | Растет |
| Синтетические реакции | Подавлены | Подавлены | Подавлены | Подавлены |
| Поглощение ионов корнями | Подавлено | Подавлено | Подавлено | Подавлено |
| Транспорт веществ | Подавлен | Подавлен | Подавлен | Подавлен |
| Концентрация пигментов | Падает | Падает | Падает | Падает |
| Деление клеток | Тормозится | Тормозится | — | — |
| Растяжение клеток | Подавлено | Подавлено | — | — |
| Число плодоэлементов | Снижено | Снижено | Снижено | Снижено |
| Старение органов | Ускорено | Ускорено | Ускорено | — |
| Биологический урожай | Понижен | Понижен | Понижен | Понижен |
Исходя из данных таблицы видно, что устойчивость растений к нескольким факторам сопровождается однонаправленными физиологическими изменениями. Это дает основание считать, что повышение устойчивости растений к одному фактору может сопровождаться повышением устойчивости к другому. Это подтверждено экспериментами.
Опытами в Институте физиологии растений РАН (Вл. В. Кузнецов и др.) показано, что кратковременная тепловая обработка растений хлопчатника сопровождается повышением их устойчивости к последующему засолению. А адаптация растений к засолению приводит к повышению их устойчивости к высокой температуре. Тепловой шок повышает способность растений приспосабливаться к последующей засухе и, наоборот, в процессе засухи повышается устойчивость организма к высокой температуре. Кратковременное воздействие высокой температурой повышает устойчивость к тяжелым металлам и УФ-Б облучению. Предшествующая засуха способствует выживанию растений в условиях засоления или холода.
Процесс повышения устойчивости организма к данному экологическому фактору в результате адаптации к фактору иной природы называется кросс-адаптацией .
Для изучения общих (неспецифических) механизмов устойчивости большой интерес представляет ответ растений на факторы, вызывающие у растений водный дефицит: на засоление, засуху, низкие и высокие температуры и некоторые другие. На уровне целого организма все растения реагируют на водный дефицит одинаково. Характерно угнетение роста побегов, усиление роста корневой системы, синтеза абсцизовой кислоты, снижение устьичной проводимости. Спустя некоторое время, ускоренно стареют нижние листья, и наблюдается их гибель. Все эти реакции направлены на снижение расходования воды за счет сокращения испаряющей поверхности, а также за счет увеличения поглотительной деятельности корня.
Специфические реакции — это реакции на действие какого-либо одного стрессового фактора. Так, фитоалексины (вещества со свойствами антибиотиков) синтезируются в растениях в ответ на контакт с болезнетворными микроорганизмами (патогенами).
Специфичность или не специфичность ответных реакций, подразумевает, с одной стороны, отношение растения к различным стрессорам и, с другой стороны, характерность реакций растений различных видов и сортов на один и тот же стрессор.
Проявление специфических и неспецифических ответных реакций растений зависит от силы стресса и скорости его развития. Специфические ответные реакции возникают чаще, если стресс развивается медленно, и организм успевает перестроиться и приспособиться к нему. Неспецифические реакции обычно возникают при более кратковременном и сильном действии стрессора. Функционирование неспецифических (общих) механизмов устойчивости позволяет растению избегать больших затрат энергии для формирования специализированных (специфических) механизмов адаптации в ответ на любое отклонение от нормы условий их обитания.
Устойчивость растений к стрессовому воздействию зависит от фазы онтогенеза. Наиболее устойчивы растения и органы растений в покоящемся состоянии: в виде семян, луковиц; древесные многолетние — в состоянии глубокого покоя после листопада. Наиболее чувствительны растения в молодом возрасте, так как в условиях стресса процессы роста повреждаются в первую очередь. Вторым критическим периодом является период формирования гамет и оплодотворения. Действие стресса в этот период приводит к снижению репродуктивной функции растений и снижению урожая.
Если стрессовые условия повторяются и имеют небольшую интенсивность, то они способствуют закаливанию растений. На этом основаны методы повышения устойчивости к низким температурам, жаре, засолению, повышенному содержанию в воздухе вредных газов.
Надежность растительного организма определяется его способностью не допускать или ликвидировать сбои на разных уровнях биологической организации: молекулярном, субклеточном, клеточном, тканевом, органном, организменном и популяционном.
Для предотвращения сбоев в жизнедеятельности растений под влиянием неблагоприятных факторов используются принципы избыточности , гетерогенности функционально равнозначных компонентов , системы репарации утраченных структур .
Избыточность структур и функциональных возможностей — один из основных способов обеспечения надежности систем. Избыточность и резервирование имеет многообразные проявления. На субклеточном уровне повышению надежности растительного организма способствуют резервирование и дублирование генетического материала. Это обеспечивается, например, двойной спиралью ДНК, увеличением плоидности. Надежность функционирования растительного организма в изменяющихся условиях поддерживается также благодаря наличию разнообразных молекул информационной РНК и образованию гетерогенных полипептидов. К ним относятся и изоферменты, которые катализируют одну и ту же реакцию, но отличаются по свои физико-химическим свойствам и устойчивостью структуры молекул в изменяющихся условиях среды.
На уровне клетки пример резервирования — избыток клеточных органелл. Так, установлено, что для обеспечения растения продуктами фотосинтеза достаточно части имеющихся хлоропластов. Остальные хлоропласты как бы остаются в резерве. То же касается и общего содержания хлорофилла. Избыточность проявляется также в большом накоплении предшественников для биосинтеза многих соединений.
На организменном уровне принцип избыточности выражается в образовании и в разновременной закладке большего, чем требуется для смены поколений, числа побегов, цветков, колосков, в огромном количестве пыльцы, семязачатков, семян.
На популяционном уровне принцип избыточности проявляется в большом числе особей, различающихся по устойчивости к тому или иному стрессовому фактору.
Системы репарации также работают на разных уровнях — молекулярном, клеточном, организменном, популяционном и биоценотическом. Репаративные процессы идут с затратой энергии и пластических веществ, поэтому репарация возможна только при сохранении достаточной интенсивности обмена веществ. Если обмен веществ прекращается, то прекращается и репарация. В экстремальных условиях внешней среды особенно большое значение имеет сохранение дыхания, так как именно дыхание обеспечивает энергией репарационные процессы.
Восстановительная способность клеток адаптированных организмов определяется устойчивостью их белков к денатурации, а именно устойчивостью связей, которые определяют вторичную, третичную и четвертичную структуру белка. Например, устойчивость зрелых семян к высоким температурам, как правило, связана с тем, что после обезвоживания их белки приобретают устойчивость к денатурации.
Главным источником энергетического материала как субстрата дыхания является фотосинтез, поэтому от устойчивости и способности фотосинтетического аппарата восстанавливаться после повреждений зависит энергообеспечение клетки и связанные с ним репарационные процессы. Для поддержания фотосинтеза в экстремальных условиях в растениях активизируется синтез компонентов мембран тилакоидов, происходит торможение окисления липидов, восстанавливается ультраструктура пластид.
На организменном уровне примером регенерации может служить развитие замещающих побегов, пробуждение спящих почек при повреждении точек роста.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
(по Н. Грину и др., 1993)
| Адаптация | Примеры | ||
| Уменьшение потери воды | |||
| Листья превращены в иглы или колючки Погруженные устьица Листья свернуты в цилиндр Толстая восковая кутикула Толстый стебель с большим отношением объема к поверхности Опушенные листья Сбрасывание листьев при засухе Устьица открыты ночью и закрыты днем Эффективная фиксация СО 2 ночью при не полностью открытых устьицах Выделение азота в виде мочевой кислоты Удлиненная петля Генле в почках Ткани выносливы к высоким температурам из-за уменьшения потоотделения или транспирации Животные прячутся в норах Дыхательные отверстия прикрыты клапанами | Cactaceae, Euphorbiaceae (молочаи), хвойные деревья Pinus, Ammophila Ammophila Листья большинства ксерофитов, насекомые Cactaceae, Euphorbiaceae («суккуленты») Многие альпийские растения Fouguieria splendens Crassulaceae (толстенковые) С-4 - растения, например Zea mays Насекомые, птицы и некоторые рептилии Пустынные млекопитающие, например верблюд, пустынная крыса Многие пустынные растения, верблюд Многие мелкие пустынные млекопитающие, например пустынная крыса Многие насекомые | ||
| Увеличение поглощения воды | |||
| Обширная поверхностная корневая система и глубоко проникающие корни Длинные корни Прорытие ходов к воде | Некоторые Cactaceae, например Opuntia и Euphorbiaceae Многие альпийские растения, например эдельвейс (Leontopodium alpinum) Термиты | ||
| Запасание воды | |||
| В слизистых клетках и в клеточных стенках В специализированны мочевом пузыре В виде жира (вода - продукт окисления) | Cactaceae и Euphorbiaceae Пустынная лягушка Пустынная крыса | ||
| Физиологическая устойчивость к потере воды | |||
| При видимом обезвоживании сохраняется жизнеспособность Потеря значительной части массы тела и быстрое ее восстановление при наличии доступной воды | Некоторые эпифитные папоротники и плауны, многие мохообразные и лишайники, осока Сагех physoides Lumbricus terrestris (теряет до 70% массы), верблюд (теряет до 30%) | ||
Окончане табл. 4.9
Совместное действие температуры
И влажности
Рассмотрение отдельных факторов среды - это не конечная цель экологического исследования, а способ подойти к сложным экологическим проблемам, дать сравнительную оценку важности различных факторов, действующих совместно в реальных экосистемах.
Температура и влажность являются ведущими климатическими факторами и тесно взаимосвязаны между собой (рис. 4.19).
Рис. 4.19. Влияние температуры на относительную влажность
воздуха (по Б. Небелу, 1993)
При неизменном количестве воды в воздухе относительная влажность увеличивается, когда температура падает. Если воздух охлаждается до температуры ниже точки водонасыщения (100%), происходит конденсация и выпадают осадки. При нагревании его относительная влажность падает. Сочетание температуры и влажности часто играет решающую роль в распределении растительности и животных. Взаимодействие температуры и влажности зависит не только от относительной, но и от абсолютной их величины. Например, температура оказывает более выраженное влияние на организмы в условиях влажности, близкой к критической, т. е. если влажность очень велика или очень мала. Влажность также играет более критическую роль при температуре, близкой к предельным значениям. Отсюда одни и те же виды организмов в различных географических зонах предпочитают разные местообитания. Так, по правилу предварения, установленному В. В. Алехиным (1951) для растительности, широко распространенные виды на юге произрастают на северных склонах, а на севере встречаются только на южных (рис. 4.20).
Рис. 4.20. Схема правила предварения (по В. В. Алехину, 1951):
1 - северный вид, обитающий на плакоре, на юге переходящий на склоны северной экспозиции и в балки; 2 - южный вид, на севере встречающийся на наиболее прогреваемых склонах южной экспозиции
Для животных выявлены принципы смены местообитания (Г. Я. Бей-Биенко, 1961) и принцип смены ярусов (М. С. Гиляров, 1970), где мезофильные виды в центре ареала, на севере его выбирают более сухие, а на юге - более влажные места или переходят от наземного образа жизни к подземному, как многие насекомые-фитофаги. Чем слабее проявляется влияние климата в конкретных местообитаниях, которые выбирает вид, тем больше их способность обитать в разных климатических условиях. Вид выбирает сочетание факторов, наиболее соответствующих его экологической валентности, путем смены местообитания, и таким образом преодолевает климатические рубежи.
Взаимосвязь температуры и влажности хорошо отражают кли-мадиаграммы, составленные по способу Вальтера-Госсена, на которых в определенных масштабах сопоставлен годовой ход температуры воздуха с ходом выпадения осадков (рис. 4.21).

Рис. 4.21. Климадиаграмма по Вальтеру-Госсену для Одессы
(по Г. Вальтеру, 1968):
а - высота над уровнем моря; б - число лет наблюдений за температурой (первая цифра) и осадками (вторая цифра); в - средняя годовая температура; г - средняя годовая сумма осадков в мм: д - средний суточный минимум самого холодного месяца; е - абсолютный минимум; ж - средний суточный максимум самого теплого месяца; з - абсолютный максимум; и - кривая средних месячных температур; к - кривая средних месячных сумм осадков (соотношение 10°=20 мм); л - то же (соотношение 10°=30 мм); м - засушливый период; н - полузасушливый период; о - влажное время года; п - месяцы со средним суточным минимумом температуры ниже 0°С; р - месяцы с абсолютным минимумом температуры ниже 0°С, с - безморозный период. По оси абсцисс - месяцы
Климадиаграммы можно построить для отдельных лет, а расположив последовательно и непрерывно одну за другой, получить климатограмму. На климатограммах легко прослеживаются экстремально сухие или экстремально холодные годы, что является весьма полезным для определения пригодности комбинаций температуры и влажности в районах предполагаемой интродукции растений или промысловых диких животных.
Атмосфера
Как уже было отмечено ранее, наша планета Земля отличается от других планет наличием воздушной оболочки, атмосферы, атмосферного воздуха. Атмосферный воздух - смесь различных газов. В его составе 78,08% азота, 20,9% кислорода, 0,93% аргона, 0,03% углекислого газа, других газов (гелий, метан, неон, ксенон, родон и др.) около 0,01%.
Значение атмосферного воздуха для живых организмов огромно и разнообразно. Это источник кислорода для дыхания и углекислоты для фотосинтеза. Он защищает живые организмы от вредных космических излучений, способствует сохранению тепла на Земле.
Атмосфера - важная часть экосферы, с которой она связана биогеохимическими циклами, включающими газообразные компоненты. Это такие, как круговороты углерода, азота, кислорода и воды. Большое значение имеют и физические свойства атмосферы. Так, воздух оказывает лишь незначительное сопротивление движению и не может служить опорой для наземных организмов, что непосредственно сказалось на их строении. Вместе с тем некоторые группы животных стали использовать полет как способ передвижения. Особо следует отметить, что в атмосфере постоянно происходит циркуляция воздушных масс, энергию которой поставляет Солнце (рис. 4.22).

Рис. 4.22. Упрощенная схема общей циркуляции
воздушных масс атмосферы:
1 – теплый воздух; 2 – охлажденный воздух; 3 – зоны высокого давления; СЕ – пассаты; СД – доминирующие юго-западные ветры; GH – полярные северо-восточные ветры
Результатом циркуляции является перераспределение водяных паров, так как атмосфера захватывает их в одном месте (где вода испаряется), переносит и отдает в другом месте (где выпадают осадки). Если же в атмосферу поступают газы, в том числе загрязняющие, такие, как двуокись серы в промышленных районах, то система атмосферной циркуляции перераспределит их и они выпадут в других местах, растворенные в дождевой воде (рис. 4.23).
Ветер, взаимодействуя с другими факторами окружающей среды, может оказывать влияние на развитие растительности, в первую очередь на деревья, растущие на открытых местах. Обычно это приводит к задержке их роста и искривлению с наветренной стороны.
Ветер играет важную роль в распространении спор, семян и т. п., расширяя возможности распространения неподвижных организмов - растений, грибов и некоторых бактерий. Ветер может оказывать влияние и на миграцию летающих животных.

Рис. 4.23. Гидрологический цикл и накопление воды
(по Е. А. Криксунову и др., 1995)
Еще одна особенность атмосферы - это ее давление, которое уменьшается с высотой. Эволюция живых организмов на нашей планете происходила при атмосферном давлении 760 мм ртутного столба на уровне моря, и оно считается «нормальным». С увеличением высоты, например при восхождении людей в горы, от недостаточной насыщенности крови кислородом может наступить состояние гипоксии или аноксии. Возникает оно вследствие того, что с возрастанием высоты над уровнем моря парциальное давление кислорода, так же как и других газов, содержащихся в атмосферном воздухе, падает. На высоте 5450 м атмосферное давление в два раза меньше, чем на уровне моря. И хотя воздух содержит здесь столько же процентов кислорода, концентрация его на единицу объема вдвое меньше.
У растений в этих условиях возрастает транспирация, что потребовало выработки адаптации для сохранения воды, как, например, у многих альпийских растений.
Топография
Топография (рельеф) относится к орографическим факторам и тесно связана с другими абиотическими факторами, хотя и не принадлежащими к таким прямодействующим экологическим факторам, как свет, тепло, вода и почва. Главным топографическим (орографическим) фактором является высота. С высотой снижаются средние температуры, увеличивается суточный перепад температур, возрастают количество осадков, скорость ветра и интенсивность радиации, понижаются атмосферное давление и концентрация газов. Так, повышение уровня местности на каждые 100 м сопровождается уменьшением температуры воздуха примерно на 0,6°С.
В зависимости от величины форм топографию или рельеф подразделяют на несколько порядков: макрорельеф (горы, межгорные впадины, низменности), мезорельеф (холмы, овраги, гряды, карстовые воронки, степные «блюдца» и др.) и микрорельеф (мелкие западинки, неровности, приствольные повышения и др.), Все это оказывает влияние на растения и животных. В результате обычным явлением стала вертикальная зональность (рис. 4.24).

Рис. 4.24. Схема, показывающая соответствие между последовательными
вертикальными и горизонтальными растительными зонами:
1 - тропическая, зона (зона тропических лесов); 2 - умеренная зона (зона лиственных и хвойных лесов); 3 – альпийская зона (зона травянистой растительности, мхов и лишайников); 4 - полярная зона (зона снегов и льдов)
Горные цепи могут служить климатическими барьерами. Влажный воздух охлаждается, поднимаясь над горами, что приводит к выпадению большого количества осадков на наветренных склонах.
На подветренной стороне горного хребта образуется так называемая «дождевая тень», воздух здесь суше, выпадает меньше осадков, создаются пустынные условия, так как воздух, опускаясь, нагревается и вбирает в себя влагу из почвы.
Это влияет на живые организмы. Для большинства позвоночных верхняя граница жизни около 6,0 км. Снижение давления с высотой влечет за собой уменьшение обеспеченности кислородом и обезвоживание животных за счет увеличения частоты дыхания. Несколько более выносливы членистоногие (ногохвостки, клещи, пауки), которые могут встречаться на ледниках, выше границы растительности. Для высокогорных растений характерен приземистый рост. Во всех высокогорных областях земного шара преобладают низкорослые стелющиеся кустарники и кустарнички (рис. 4.25), подушковидные и розеточные многолетние травы, дерновидные злаки и осоки, мхи и лишайники.

Рис. 4.25. Можжевельник туркестанский - на склонах хребта
Терекей-Алатау (по И. Г. Серебрякову, 1955):
А – древовидная форма (лугово-лесной пояс, 2900 м над уровнем моря); Б – стланник (субальпийский пояс, 3200 м над уровнем моря)
Характерная морфологическая черта многих высокогорных приземистых растений, например кустарников и кустарничков, - значительное преобладание подземной массы по сравнению с надземной.
Низкорослость высокогорных растений связывают с адаптацией к низким температурам и с формообразующим действием радиации, богатой коротковолновой частью спектра, тормозящей ростовые процессы. В анатомическом строении высокогорных растений есть ряд черт, которые способствуют защите от избыточной радиации, связаны с характером водного режима и обмена веществ в высокогорьях: утолщение покровных тканей, придающих устойчивость к сильным ветрам и т. д. У растений, живущих на скалах, наблюдаются изменения в сторону ксероморфоза: уменьшаются размеры клеток и возрастает плотность тканей, увеличивается число устьиц на единицу поверхности листа, уменьшаются их размеры. У видов же, обитающих вблизи талых вод или других источников увлажнения, листья крупнее и ксероморфные черты выражены слабее.
Низкие температуры и сильная освещенность способствуют образованию больших количеств антоциана, отсюда глубокие, насыщенные тона окраски цветов. Сочетание небольших листьев при малом росте и крупных яркоокрашенных цветков - характерная черта многих альпийских растений.
Характерная черта физиологии и биохимии высокогорных растений - повышение интенсивности окислительно-восстановительных процессов, увеличение активности участвующих в них ферментов (каталазы, пероксидазы и др.), более низкие, чем у равнинных растений, температурные оптимумы их работы.
Дыхание высокогорных растений устойчиво к неблагоприятным воздействиям, как правило, наблюдается усиление дыхания, а следовательно, и увеличение энергии, освобождающейся при распаде сложных соединений. По современным представлениям, это является одной из физиологических основ приспособленности растений к крайним условиям.
При поднятии в горы меняется и сезонное развитие растений. Так, весной, поднимаясь в горы, можно видеть развитие одного и того же вида в следующей последовательности: в низкогорном поясе - цветение, в среднем - бутонизацию, еще выше - начало вегетации и, наконец, только появление после таяния снега. Осенью же при подъеме в горы наблюдаем ускоренное наступление осенних фенофаз: расцвечивание листвы, листопад, отмирание надземных частей. Четко прослеживается сокращение у растений вегетационного периода.
Наряду с высотой над уровнем моря большое значение для живых организмов имеют экспозиция и крутизна склонов.
В северном полушарии склоны гор, обращенные на юг, получают больше солнечного света, интенсивность света и температура здесь выше, чем на дне долины и на склонах северной экспозиции. В южном же полушарии наблюдается обратная ситуация. Это оказывает поразительное влияние как на естественную растительность, так и на угодья, используемые человеком. Например, широкие расщелины между скалами над Дунаем в восточной Сербии, защищенные от ветров и испытывающие увлажняющее действие реки, способствовали сохранению многих редких, реликтовых и эндемических видов растений, среди них «медвежий орешник» -Corylus colurna, грецкий орех - Juglans regia, сирень (дикая форма) - Syringa vulgaris и др.
Для крутых склонов характерны быстрый дренаж и смывание почв. Здесь почвы обычно маломощные и более сухие, с ксероморфной растительностью. При уклоне, превышающем 35°, почва не образуется, растительность отсутствует, создаются осыпи из рыхлого материала.
Прочие физические факторы
К прочим физическим факторам, окружающим живые организмы на Земле, относят главным образом атмосферное электричество, огонь, шум, магнитное поле Земли, ионизирующие излучения.
Атмосферное электричество действует на живые организмы посредством разрядов и ионизации воздуха. Например, известно губительное действие молний при попадании в крупные деревья, животных. Есть определенные закономерности в частоте повреждаемости молнией различных древесных пород. Это связывают как с формой кроны, так и с электропроводящими свойствами коры, например с быстротой ее намокания. По частоте поражения молниями на первом месте стоят ель и сосна, затем береза, а осина повреждается значительно реже. Молнии вызывают механическое повреждение деревьев (расщепление стволов, трещины), выпадение крупных деревьев, тем самым оказывают влияние на структуру древостоя, зачастую являются причиной возникновения пожаров. Около 21% пожаров лесных угодий России происходит по вине молний, при грозах.
Роль атмосферных электрических разрядов состоит и в том, что они во время грозы из атмосферного азота и кислорода синтезируют окиси азота, которые с дождевыми водами попадают в почву и накапливаются в ней от 4 до 10 кг в год на 1 гектар в форме селитры и азотной кислоты.
Действие ионизации воздуха на человека, животных и растения еще недостаточно изучено. Вместе с тем достоверно установлена прямая зависимость между самочувствием человека и присутствием легких ионов в воздухе. Высказывается мнение, что ионизация воздуха служит материальной способности некоторых растений «предсказывать погоду» (снижение фотосинтеза и дыхания, закрывание устьиц и прекращение транспирации перед грозой задолго до падения атмосферного давления). Экспериментально доказано влияние слабого тока на корневые системы некоторых растений. Например, у саженцев ели и сосны фитомасса увеличивается на 100-120%. Установлена возможность с помощью воздействия направленного электрического поля регулировать темпы перемещения веществ внутри дерева, а следовательно, и темпы его роста.
Огонь в жизни растений и животных - довольно редкий, но весьма действенный фактор. Пожары, например, в лесах, как уже было отмечено ранее, могут возникать как естественным путем от ударов молний, так и по вине человека, его деятельности. Поэтому огонь относят как к естественным экологическим факторам, так и антропогенным.
Серьезные последствия имеют не только верховые лесные пожары, охватывающие весь древостой, но и низовые, которые губят напочвенную растительность, подрост, нижние ветви деревьев, нередко корневую систему. Гибнут животные. Кроме повреждений непосредственно от огня пожары вызывают ухудшение состояния древостоя. Снижается прирост. Ослабленные деревья в большей степени заражаются грибами, такими, как древесная гниль, легко проникающими через «огневые раны», подвергаются нападению насекомых-вредителей.
Лесные пожары сильно изменяют условия обитания растений и животных. Во время пожара в хвойных лесах температура доходит до 800-900°С, в почве на глубине 3,5 см - до 95°С, на глубине 7см - до 70"С. В сухих лесах практически полностью сгорает подстилка и почвенный гумус. Минеральные частицы верхнего слоя почвы спекаются. Образуются комки или стекловидная корка, трудно проницаемые для воздуха, воды и корней. Почва сильно уплотняется. От сгорания органических кислот и освобождения оснований кислотность почвы резко уменьшается, в верхних горизонтах значение рН нередко доходит до сильнощелочного. От высокой температуры верхние слои почвы стерилизуются - гибнет почвенная микрофлора, а в более глубоких - изменяется ее состав, происходит обеднение наиболее важными для жизнедеятельности растений группами. Так, в почвах хвойных лесов после пожаров преобладает деятельность микроорганизмов, вызывающих масляно-кислое брожение и денитрификацию.
После лесных пожаров происходит резкое изменение условий в растительных сообществах (осветление, изменение температурного и других факторов микроклимата), особенно когда произошло уничтожение древостоя, и ведет к тому, что в дальнейшем гари заселяются видами живых организмов с различными адаптивными особенностями, помогающими перенести пожар и выжить на гарях. Так, у растений это глубокие подземные почки возобновления, способность семян долго сохраняться в почве и выдерживать высокую температуру, выносливость к заморозкам, сильной освещенности и т. д.
Возобновление растительности на гарях имеет свои особенности. На выжженных местах из спор, занесенных ветром, появляются мхи-пионеры, через три - пять лет из мхов наиболее обилен «пожарный мох» - Funaria hygrometrica. Из высших растений быстро заселяет гари иван-чай (Chamaenerion angustifolion). Постепенное заселение гарей происходит и древесной растительностью - ивой, березой, осиной и др. (рис. 4.26).

Рис. 4.26. Влияние пожара на растительность древесных «колков»
Зауральской лесостепи (по Д. Ф. Федюнину, 1953) :
А - до пожара; Б - после пожара; В - через год после пожара; 1 - ива; 2 - береза, 3 - осина
Степные пожары («палы») могут быть более или менее регулярными, связанными с деятельностью человека, и играть существенную роль в жизни живых организмов, иногда и положительную для регулирования роста, возобновления, отбора видов и поддержания постоянного состава травостоя.
Шум как естественный экологический фактор для живых организмов несуществен, но может оказывать и существенное воздействие с усилением антропогенных воздействий (шум, возникающий при работе транспортных средств, оборудования промышленных и бытовых предприятий, вентиляционных и газотурбинных установок и др.).
Величину звуковых давлений изменяют и нормируют в децибелах. Весь диапазон слышимых человеком звуков укладывается в 150 дБ. На нашей планете жизнь организмов проходит в мире звуков. Например, орган слуха человека приспособлен к некоторым постоянным или повторяющимся шумам (слуховая адаптация). Человек теряет работоспособность без привычных шумов. Сильный шум еще более отрицательно сказывается на здоровье человека. У людей, живущих и работающих в неблагоприятных акустических условиях, имеются признаки изменения функционального состояния центральной нервной и сердечно-сосудистой систем.
Исследованиями доказано воздействие шума и на растительные организмы. Так, растения близ аэродромов, с которых непрерывно стартуют реактивные самолеты, испытывают угнетение роста и даже отмечается исчезновение отдельных видов. В целом ряде научных работ показано угнетающее действие шума (около 100 дБ с частотой звука от 31,5 до 90 тыс. Гц) на растения табака, где обнаруживали снижение интенсивности роста листьев, в первую очередь у молодых растений. Привлекает внимание ученых и действие ритмических звуков на растения. Исследования по изучению действия музыки на растения (кукуруза, тыква, петуния, циния, календула), проведенные в 1969 г. американским музыкантом и певицей Д. Ретолэк, показали, что на музыку Баха и индийские музыкальные мелодии растения отзывались положительно. Их габитус, сухой вес биомассы были наибольшими по сравнению с контролем. И что самое удивительное, так это то, что их стебли прямо-таки тянулись к источнику этих звуков. В то же время на рок-музыку и непрерывные барабанные ритмы зеленые растения отвечали уменьшением размеров листьев и корней, снижением массы, и все они отклонялись от источника звука, как будто бы хотели уйти от губительного действия музыки (рис. 4.27).

Рис. 4.27. Вид растений после действия разной музыки:
А - индийские мелодии (Р. Шанкар); Б - музыка И.-С. Баха; В - рок-музыка (опыты Д. Ретолэк, 1969)
Растения, подобно людям, реагируют на музыку как целостный живой организм. Их чувствительными «нервными» проводниками, по мнению ряда ученых, являются флоэмные пучки, меристема и возбудимые клетки, расположенные в разных частях растения, связанные между собой биоэлектрическими процессами. Вероятно, этот факт - одна из причин сходства реакции на музыку у растений, животных и человека.
Магнитное поле Земли. Наша планета Земля обладает магнитными свойствами. Стрелка компаса всегда ориентируется по магнитному меридиану, указывая одним концом на север, другим - на юг. МагнитологГпоказали, что для создания наблюдаемого геомагнитного поля в центре Земли необходимо поместить гигантский цилиндрический магнит диаметром 200 км и длиной 4000 км. Ось земного магнита расположена под углом 1,5" к оси вращения Земли, поэтому магнитные полюса не совпадают с географическими. Со временем магнитные полюса меняют свое положение. Установлено, что северный магнитный полюс за сутки перемещается по поверхности Земли на 20,5 м, или 7,5 км в год, а Южный - на 30 м (11 км в год). Как у всякого магнита, магнитные силовые линии Земли выходят из одного полюса и через околоземное пространство замыкаются в другом полюсе. За счет этого явления около Земли создается магнитосфера (рис. 4.28).

Рис. 4.28. Меридиональные сечения магнитосферы Земли:
1 - солнечный ветер; 2 - ударный фронт; 3 - магнитная полость; 4 - магнитопауза; 5 - верхняя граница магнитосферной щели; 6 - плазменная мантия; 7 - внешний радиационный пояс; 8 - внутренний радиационный пояс, или плазмосфера; 9 - нейтральный слой; 10 - плазменный слой
Она задерживает потоки солнечных заряженных частиц, называемых плазмой, или солнечным ветром, не пропуская их к поверхности планеты. Солнечный ветер как бы огибает Землю и смещается на ночную сторону, вытягивая, в свою очередь, и магнитные силовые линии в этом же направлении. Деформация магнитных силовых линий связана с тем, что потоки солнечной плазмы несут с собой как бы «вмороженное» магнитное поле, которое и взаимодействует с магнитосферой Земли. За последние 600 тыс. лет палеомагнитологи зафиксировали 12 эпох инверсии геомагнитного поля (табл. 4.10).